| THE NERVOUS SYSTEM is the most complicated and highly organized of the various systems which make up the human body. It is the mechanism concerned with the correlation and integration of various bodily processes and the reactions and adjustments of the organism to its environment. In addition the cerebral cortex is concerned with conscious life. It may be divided into two parts, central and peripheral. | 1 |
| The central nervous system consists of the encephalon or brain, contained within the cranium, and the medulla spinalis or spinal cord, lodged in the vertebral canal; the two portions are continuous with one another at the level of the upper border of the atlas vertebra. | 2 |
| The peripheral nervous system consists of a series of nerves by which the central nervous system is connected with the various tissues of the body. For descriptive purposes these nerves may be arranged in two groups, cerebrospinal and sympathetic, the arrangement, however, being an arbitrary one, since the two groups are intimately connected and closely intermingled. Both the cerebrospinal and sympathetic nerves have nuclei of origin (the somatic efferent and sympathetic efferent) as well as nuclei of termination (somatic afferent and sympathetic afferent) in the central nervous system. The cerebrospinal nerves are forty-three in number on either side—twelve cranial, attached to the brain, and thirty-one spinal, to the medulla spinalis. They are associated with the functions of the special and general senses and with the voluntary movements of the body. The sympathetic nerves transmit the impulses which regulate the movements of the viscera, determine the caliber of the bloodvessels, and control the phenomena of secretion. In relation with them are two rows of central ganglia, situated one on either side of the middle line in front of the vertebral column; these ganglia are intimately connected with the medulla spinalis and the spinal nerves, and are also joined to each other by vertical strands of nerve fibers so as to constitute a pair of knotted cords, the sympathetic trunks, which reach from the base of the skull to the coccyx. The sympathetic nerves issuing from the ganglia form three great prevertebral plexuses which supply the thoracic, abdominal, and pelvic viscera; in relation to the walls of these viscera intricate nerve plexuses and numerous peripheral ganglia are found. | 3 |
| |
| 1. Structure of the Nervous System |
| |
| The nervous tissues are composed of nerve cells and their various processes, together with a supporting tissue called neuroglia, which, however, is found only in the brain and medulla spinalis. Certain long processes of the nerve cells are of special importance, and it is convenient to consider them apart from the cells; they are known as nerve fibers. | 4 |
| To the naked eye a difference is obvious between certain portions of the brain and medulla spinalis, viz., the gray substance and the white substance. The gray substance is largely composed of nerve cells, while the white substance contains only their long processes, the nerve fibers. It is in the former that nervous impressions are received, stored, and transformed into efferent impulses, and by the latter that they are conducted. Hence the gray substance forms the essential constituent of all the ganglionic centers, both those in the isolated ganglia and those aggregated in the brain and medulla spinalis; while the white substance forms the bulk of the commissural portions of the nerve centers and the peripheral nerves. | 5 |
| |
| Neuroglia.—Neuroglia, the peculiar ground substance in which are imbedded the true nervous constituents of the brain and medulla spinalis, consists of cells and fibers. Some of the cells are stellate in shape, with ill-defined cell body, and their fine processes become neuroglia fibers, which extend radially and unbranched (Fig. 623, B) among the nerve cells and fibers which they aid in supporting. Other cells give off fibers which branch repeatedly (Fig. 623, A). Some of the fibers start from the epithelial cells lining the ventricles of the brain and central canal of the medulla spinalis, and pass through the nervous tissue, branching repeatedly to end in slight enlargements on the pia mater. Thus, neuroglia is evidently a connective tissue in function but is not so in development; it is ectodermal in origin, whereas all connective tissues are mesodermal. | 6 |
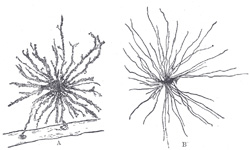 |
FIG. 623– Neuroglia cells of brain shown by Golgi’s method. A. Cell with branched processes. B. Spider cell with unbranched processes. (After Andriezen.) (See enlarged image) |
| |
| |
| Nerve Cells (Fig. 624).—Nerve cells are largely aggregated in the gray substance of the brain and medulla spinalis, but smaller collections of these cells also form the swellings, called ganglia, seen on many nerves. These latter are found chiefly upon the spinal and cranial nerve roots and in connection with the sympathetic nerves. | 7 |
| The nerve cells vary in shape and size, and have one or more processes. They may be divided for purposes of description into three groups, according to the number of processes which they possess: (1) Unipolar cells, which are found in the spinal ganglia; the single process, after a short course, divides in a T-shaped manner (Fig. 624, E). (2) Bipolar cells, also found in the spinal ganglia (Fig. 625), when the cells are in an embryonic condition. They are best demonstrated in the spinal ganglia of fish. Sometimes the processes come off from opposite poles of the cell, and the cell then assumes a spindle shape; in other cells both processes emerge at the same point. In some cases where two fibers are apparently connected with a cell, one of the fibers is really derived from an adjoining nerve cell and is passing to end in a ramification around the ganglion cell, or, again, it may be coiled spirally around the nerve process which is issuing from the cell. (3) Multipolar cells, which are pyramidal or stellate in shape, and characterized by their large size and by the numerous processes which issue from them. The processes are of two kinds: one of them is termed the axis-cylinder process or axon because it becomes the axis-cylinder of a nerve fiber (Figs. 626, 627, 628). The others are termed the protoplasmic processes or dendrons; they begin to divide and subdivide soon after they emerge from the cell, and finally end in minute twigs and become lost among the other elements of the nervous tissue. | 8 |
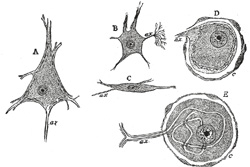 |
FIG. 624– Various forms of nerve cells. A. Pyramidal cell. B. Small multipolar cell, in which the axon quickly divides into numerous branches. C. Small fusiform cell. D and E. Ganglion cells (E shows T-shaped division of axon). ax. Axon. c. Capsule. (See enlarged image) |
| |
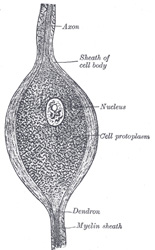 |
FIG. 625– Bipolar nerve cell from the spinal ganglion of the pike. (After Kölliker.) (See enlarged image) |
| |
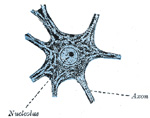 |
FIG. 626– Motor nerve cell from ventral horn of medulla spinalis of rabbit. The angular and spindle-shaped Nissl bodies are well shown. (After Nissl.) (See enlarged image) |
| |
| The body of the nerve cell, known as the cyton, consists of a finely fibrillated protoplasmic material, of a reddish or yellowishbrown color, which occasionally presents patches of a deeper tint, caused by the aggregation of pigment granules at one side of the nucleus, as in the substantia nigra and locus cæruleus of the brain. The protoplasm also contains peculiar angular granules, which stain deeply with basic dyes, such as methylene blue; these are known as Nissl’s granules (Fig. 626). They extend into the dendritic processes but not into the axis-cylinder; the small clear area at the point of exit of the axon in some cell types is termed the cone of origin. These granules disappear (chromatolysis) during fatigue or after prolonged stimulation of the nerve fibers connected with the cells. They are supposed to represent a store of nervous energy, and in various mental diseases are deficient or absent. The nucleus is, as a rule, a large, well-defined, spherical body, often presenting an intranuclear network, and containing a well-marked nucleolus. | 9 |
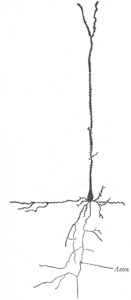 |
FIG. 627– Pyramidal cell from the cerebral cortex of a mouse. (After Ramón y Cajal.) (See enlarged image) |
| |
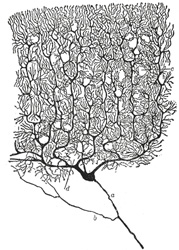 |
FIG. 628– Cell of Purkinje from the cerebellum. Golgi method. (Cajal.) a. Axon. b. Collateral. c and d. Dendrons. (See enlarged image) |
| |
| In addition to the protoplasmic network described above, each nerve cell may be shown to have delicate neurofibrils running through its substance (Fig. 629); these fibrils are continuous with the fibrils of the axon, and are believed to convey nerve impulses. Golgi has also described an extracellular network, which is probably a supporting structure. | 10 |
| |
| Nerve Fibers.—Nerve fibers are found universally in the peripheral nerves and in the white substance of the brain and medulla spinalis. They are of two kinds—viz., medullated or white fibers, and non-medullated or gray fibers. | 11 |
| The medullated fibers form the white part of the brain and medulla spinalis, and also the greater part of every cranial and spinal nerve, and give to these structures their opaque, white aspect. When perfectly fresh they appear to be homogeneous; but soon after removal from the body each fiber presents, when examined by transmitted light, a double outline or contour, as if consisting of two parts (Fig. 630). The central portion is named the axis-cylinder; around this is a sheath of fatty material, staining black with osmic acid, named the white substance of Schwann or medullary sheath, which gives to the fiber its double contour, and the whole is enclosed in a delicate membrane, the neurolemma, primitive sheath, or nucleated sheath of Schwann (Fig. 633) | 12 |
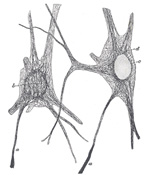 |
FIG. 629– Nerve cells of kitten, showing neurofibrils. (Cajal.) a. Axon. b. Cyton. c. Nucleus. d. Neurofibrils. (See enlarged image) |
| |
| The axis-cylinder is the essential part of the nerve fiber, and is always present; the medullary sheath and the neurolemma are occasionally absent, expecially at the origin and termination of the nerve fiber. The axis-cylinder undergoes no interruption from its origin in the nerve center to its peripheral termination, and must be regarded as a direct prolongation of a nerve cell. It constitutes about one-half or one-third of the nerve fiber, being greater in proportion in the fibers of the central organs than in those of the nerves. It is quite transparent, and is therefore indistinguishable in a perfectly fresh and natural state of the nerve. It is made up of exceedingly fine fibrils, which stain darkly with gold chloride (Fig. 632), and at its termination may be seen to break up into these fibrillæ. The fibrillæ have been termed the primitive fibrillæ of Schultze. The axis-cylinder is said by some to be enveloped in a special reticular sheath, which separates it from the medullary sheath, and is composed of a substance called neurokeratin. The more common opinion is that this network or reticulum is contained in the white matter of Schwann, and by some it is believed to be produced by the action of the reagents employed to show it. | 13 |
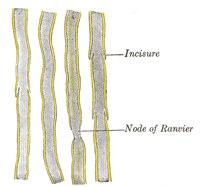 |
FIG. 630– Medullated nerve fibers. X 350. (See enlarged image) |
| |
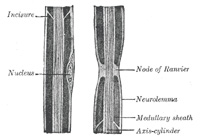 |
FIG. 631– Diagram of longitudinal sections of medullated nerve fibers. Osmic acid. (See enlarged image) |
| |
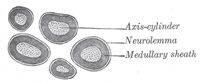 |
FIG. 632– Transverse sections of medullated nerve fibers. Osmic acid. (See enlarged image) |
| |
 |
FIG. 633– Diagram of medullated nerve fibers stained with osmic acid. X 425. (Schäfer.) R. Nodes of Ranvier. a. Neurolemma. c. Nucleus. (See enlarged image) |
| |
| The medullary sheath, or white matter of Schwann (Fig. 631), is regarded as being a fatty matter in a fluid state, which insulates and protects the essential part of the nerve—the axis-cylinder. It varies in thickness, in some forming a layer of extreme thinness, so as to be scarcely distinguishable, in others forming about one-half the nerve fiber. The variation in diameter of the nerve fibers (from 2 to 16μ) depends mainly upon the amount of the white substance, though the axis cylinder also varies within certain limits. The medullary sheath undergoes interruptions in its continuity at regular intervals, giving to the fiber the appearance of constriction at these points: these are known as the nodes of Ranvier (Figs. 631 and 633). The portion of nerve fiber between two nodes is called an internodal segment. The neurolemma or primitive sheath is not interrupted at the nodes, but passes over them as a continuous membrane. If the fiber be treated with silver nitrate the reagent penetrates the neurolemma at the nodes, and on exposure to light reduction takes place, giving rise to the appearance of black crosses, Ranvier’s crosses, on the axis-cylinder. There may also be seen transverse lines beyond the nodes termed Frommann’s lines (Fig. 634); the significance of these is not understood. In addition to these interruptions oblique clefts may be seen in the medullary sheath, subdividing it into irregular portions, which are termed medullary segments, or segments of Lantermann (Fig. 631); there is reason to believe that these clefts are artificially produced in the preparation of the specimens. Medullated nerve fibers, when examined in the fresh condition, frequently present a beaded or varicose appearance: this is due to manipulation and pressure causing the oily matter to collect into drops, and in consequence of the extreme delicacy of the primitive sheath, even slight pressure will cause the transudation of the fatty matter, which collects as drops of oil outside the membrane. | 14 |
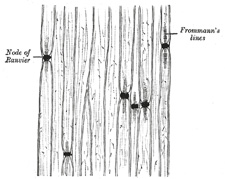 |
FIG. 634– Medullated nerve fibers stained with silver nitrate. (See enlarged image) |
| |
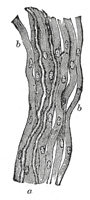 |
FIG. 635– A small nervous branch from the sympathetic of a mammal. a. Two medullated nerve fibers among a number of gray nerve fibers, b. (See enlarged image) |
| |
| The neurolemma or primitive sheath presents the appearance of a delicate, structureless membrane. Here and there beneath it, and situated in depressions in the white matter of Schwann, are nuclei surrounded by a small amount of protoplasm. The nuclei are oval and somewhat flattened, and bear a definite relation to the nodes of Ranvier, one nucleus generally lying in the center of each internode. The primitive sheath is not present in all medullated nerve fibers, being absent in those fibers which are found in the brain and medulla spinalis. | 15 |
| |
| Wallerian Degeneration.—When nerve fibers are cut across, the central ends of the fibers degenerate as far as the first node of Ranvier; but the peripheral ends degenerate simultaneously throughout their whole length. The axons break up into fragments and become surrounded by drops of fatty substance which are formed from the breaking down of the medullary sheath. The nuclei of the primitive sheath proliferate, and finally absorption of the axons and fatty substance occurs. If the cut ends of the nerve be sutured together regeneration of the nerve fibers takes place by the downgrowth of axons from the central end of the nerve. At one time it was believed that the regeneration was peripheral in origin, but this has been disproved, the proliferated nuclei in the peripheral portions taking part merely in the formation of the so-called scaffolding along which the new axons pass. | 16 |
| |
| Non-medullated Fibers.—Most of the fibers of the sympathetic system, and some of the cerebrospinal, consist of the gray or gelatinous nerve fibers (fibers of Remak) (Fig. 635). Each of these consists of an axis-cylinder to which nuclei are applied at intervals. These nuclei are believed to be in connection with a delicate sheath corresponding with the neurolemma of the medullated nerve fiber. In external appearance the non-medullated nerve fibers are semitransparent and gray or yellowish gray. The individual fibers vary in size, generally averaging about half the size of the medullated fibers. | 17 |
| |
| Structure of the Peripheral Nerves and Ganglia.—The cerebrospinal nerves consist of numerous nerve fibers collected together and enclosed in membranous sheaths (Fig. 636). A small bundle of fibers, enclosed in a tubular sheath, is called a funiculus; if the nerve is of small size, it may consist only of a single funiculus; but if large, the funiculi are collected together into larger bundles or fasciculi, which are bound together in a common membranous investment. In structure the common membranous investment, or sheath of the whole nerve (epineurium), as well as the septa given off from it to separate the fasciculi, consist of connective tissue, composed of white and yellow elastic fibers, the latter existing in great abundance. The tubular sheath of the funiculi (perineurium) is a fine, smooth, transparent membrane, which may be easily separated, in the form of a tube, from the fibers it encloses; in structure it is made up of connective tissue, which has a distinctly lamellar arrangement. The nerve fibers are held together and supported within the funiculus by delicate connective tissue, called the endoneurium. It is continuous with septa which pass inward from the innermost layer of the perineurium, and shows a ground substance in which are imbedded fine bundles of fibrous connective tissue running for the most part longitudinally. It serves to support capillary vessels, arranged so as to form a net-work with elongated meshes. The cerebrospinal nerves consist almost exclusively of medullated nerve fibers, only a very small proportion of non-medullated being present. | 18 |
| The bloodvessels supplying a nerve end in a minute capillary plexus, the vessels composing which pierce the perineurium, and run, for the most part, parallel with the fibers; they are connected together by short, transverse vessels, forming narrow, oblong meshes, similar to the capillary system of muscle. Fine non-medullated nerve fibers, vasomotor fibers, accompany these capillary vessels, and break up into elementary fibrils, which form a network around the vessels. Horsley has demonstrated certain medullated fibers running in the epineurium and terminating in small spheroidal tactile corpuscles or end bulbs of Krause. These nerve fibers, which Marshall believes to be sensory, and which he has termed nervi nervorum, are considered by him to have an important bearing upon certain neuralgic pains. | 19 |
| The nerve fibers, so far as is at present known, do not coalesce, but pursue an uninterrupted course from the center to the periphery. In separating a nerve, however, into its component funiculi, it may be seen that these do not pursue a perfectly insulated course, but occasionally join at a very acute angle with other funiculi proceeding in the same direction; from this, branches are given off, to joint again in like manner with other funiculi. It must be distinctly understood, however, that in these communications the individual nerve fibers do not coalesce, but merely pass into the sheath of the adjacent nerve, become intermixed with its nerve fibers, and again pass on to intermingle with the nerve fibers in some adjoining funiculus. | 20 |
| Nerves, in their course, subdivide into branches, and these frequently communicate with branches of a neighboring nerve. The communications which thus take place form what is called a plexus. Sometimes a plexus is formed by the primary branches of the trunks of the nerves—as the cervical, brachial, lumbar, and sacral plexuses—and occasionally by the terminal funiculi, as in the plexuses formed at the periphery of the body. In the formation of a plexus, the component nerves divide, then join, and again subdivide in such a complex manner that the individual funiculi become interlaced most intricate’y; so that each branch leaving a plexus may contain filaments from all the primary nervous trunks which form the plexus. In the formation also of smaller plexuses at the periphery of the body there is a free interchange of the funiculi and primitive fibers. In each case, however, the individual fibers remain separate and distinct. | 21 |
| It is probable that through this interchange of fibers, every branch passing off from a plexus has a more extensive connection with the spinal cord than if it had proceeded to its distribution without forming connections with other nerves. Consequently the parts supplied by these nerves have more extended relations with the nervous centers; by this means, also, groups of muscles may be associated for combined action. | 22 |
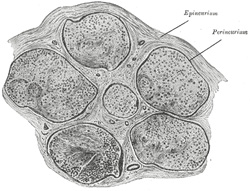 |
FIG. 636– Transverse section of human tibial nerve. (See enlarged image) |
| |
| The sympathetic nerves are constructed in the same manner as the cerebrospinal nerves, but consist mainly of non-medullated fibers, collected in funiculi and enclosed in sheaths of connective tissue. There is, however, in these nerves a certain admixture of medullated fibers. The number of the latter varies in different nerves, and may be estimated by the color of the nerve. Those branches of the sympathetic, which present a well-marked gray color, are composed chiefly of non-medullated nerve fibers, intermixed with a few medullated fibers; while those of a white color contain many of the latter fibers, and few of the former. | 23 |
| The cerebrospinal and sympathetic nerve fibers convey various impressions. The sensory nerves, called also centripetal or afferent nerves, transmit to the nervous centers impressions made upon the peripheral extremities of the nerves, and in this way the mind, through the medium of the brain, becomes conscious of external objects. The centrifugal or efferent nerves transmit impressions from the nervous centers to the parts to which the nerves are distributed, these impressions either exciting muscular contraction or influencing the processes of nutrition, growth, and secretion. | 24 |
| |
| Origins and Terminations of Nerves.—By the expression “the terminations of nerve fibers” is signified their connections with the nerve centers and with the parts they supply. The former are sometimes called their origins or central terminations; the latter their peripheral terminations. | 25 |
| |
| Origins of Nerves.—The origin in some cases is single—that is to say, the whole nerve emerges from the nervous center by a single root; in other instances the nerve arises by two or more roots which come off from different parts of the nerve center, sometimes widely apart from each other, and it often happens, when a nerve arises in this way by two roots, that the functions of these two roots are different; as, for example, in the spinal nerves, each of which arises by two roots, the anterior of which is motor, and the posterior sensory. The point where the nerve root or roots emerge from the surface of the nervous center is named the superficial or apparent origin, but the fibers of the nerve can be traced for a certain distance into the substance of the nervous center to some portion of the gray matter, which constitutes the deep or real origin of the nerve. The centrifugal or efferent nerve fibers originate in the nerve cells of the gray substance, the axis-cylinder processes of these cells being prolonged to form the fibers. In the case of the centripetal or afferent nerves the fibers grow inward either from nerve cells in the organs of special sense, e. g., the retina, or from nerve cells in the ganglia. Having entered the nerve center they branch and send their ultimate twigs among the cells, without, however, uniting with them. | 26 |
| |
| Peripheral Terminations of Nerves.—Nerve fibers terminate peripherally in various ways, and these may be conveniently studied in the sensory and motor nerves respectively. The terminations of the sensory nerves are dealt with in the section on Sense Organs. | 27 |
| Motor nerves can be traced into either unstriped or striped muscular fibers. In the unstriped or involuntary muscles the nerves are derived from the sympathetic, and are composed mainly of non-medullated fibers. Near their terminations they divide into numerous branches, which communicate and form intimate plexuses. At the junction of the branches small triangular nuclear bodies (ganglion cells) are situated. From these plexuses minute branches are given off which divide and break up into the ultimate fibrillæ of which the nerves are composed. These fibrillæ course between the involuntary muscle cells, and, according to Elischer, terminate on the surfaces of the cells, opposite the nuclei, in minute swellings. | 28 |
| In the striped or voluntary muscle the nerves supplying the muscular fibers are derived from the cerebrospinal nerves, and are composed mainly of medullated fibers. The nerve, after entering the sheath of the muscle, breaks up into fibers or bundles of fibers, which form plexuses, and gradually divide until, as a rule, a single nerve fiber enters a single muscular fiber. Sometimes, however, if the muscular fiber be long, more than one nerve fiber enters it. Within the muscular fiber the nerve terminates in a special expansion, called by Kühne, who first accurately described it, a motor end-plate (Fig. 637). The nerve fiber, on approaching the muscular fiber, suddenly loses its medullary sheath, the neurolemma becomes continuous with the sarcolemma of the muscle, and only the axis-cylinder enters the muscular fiber. There it at once spreads out, ramifying like the roots of a tree, immediately beneath the sarcolemma, and becomes imbedded in a layer of granular matter, containing a number of clear, oblong nuclei, the whole constituting an end-plate from which the contractile wave of the muscular fiber is said to start. | 29 |
| Ganglia are small aggregations of nerve cells. They are found on the posterior roots of the spinal nerves; on the sensory roots of the trigeminal, facial, glossopharyngeal, and vagus nerves, and on the acoustic nerves. They are also found in connection with the sympathetic nerves. On section they are seen to consist of a reddish-gray substance, traversed by numerous white nerve fibers; they vary considerably in form and size; the largest are found in the cavity of the abdomen; the smallest, not visible to the naked eye, exist in considerable numbers upon the nerves distributed to the different viscera. Each ganglion is invested by a smooth and firm, closely adhering, membranous envelope, consisting of dense areolar tissue; this sheath is continuous with the perineurium of the nerves, and sends numerous processes into the interior to support the bloodvessels supplying the substance of the ganglion. | 30 |
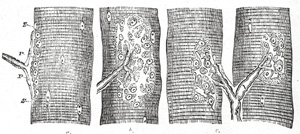 |
FIG. 637– Muscular fibers of Lacerta viridis with the terminations of nerves. a. Seen in profile. P, P. The nerve end-plates. S, S. The base of the plate, consisting of a granular mass with nuclei. b. The same as seen in looking at a perfectly fresh fiber, the nervous ends being probably still excitable. (The forms of the variously divided plate can hardly be represented in a woodcut by sufficiently delicate and pale contours to reproduce correctly what is seen in nature.) c. The same as seen two hours after death from poisoning by curare. (See enlarged image) |
| |
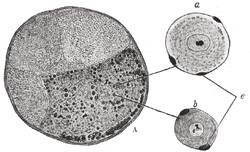 |
FIG. 638– Transverse section of spinal ganglion of rabbit. A. Ganglion. X 30. a. Large clear nerve cell. b. Small deeply staining nerve cell. c. Nuclei of capsule. X 250. The lines in the center point to the corresponding cells in the ganglion. (See enlarged image) |
| |
| In structure all ganglia are essentially similar, consisting of the same structural elements—viz., nerve cells and nerve fibers. Each nerve cell has a nucleated sheath which is continuous with the neurolemma of the nerve fiber with which the cell is connected. The nerve cells in the ganglia of the spinal nerves (Fig. 638) are pyriform in shape, and have each a single process. A short distance from the cell and while still within the ganglion this process divides in a T-shaped manner, one limb of the cross-bar turning into the medulla spinalis, the other limb passing outward to the periphery. In the sympathetic ganglia (Fig. 639) the nerve cells are multipolar and each has one axis-cylinder process and several dendrons; the axon emerges from the ganglion as a non-medullated nerve fiber. Similar cells are found in the ganglia connected with the trigeminal nerve, and these ganglia are therefore regarded as the cranial portions of the sympathetic system. The sympathetic nervous system includes those portions of the nervous mechanism in which a medullated nerve fiber from the central system passes to a ganglion, sympathetic or peripheral, from which fibers, usually non-medullated, are distributed to such structures, e. g., bloodvessels, as are not under voluntary control. The spinal and sympathetic ganglia differ somewhat in the size and disposition of the cells and in the number of nerve fibers entering and leaving them. In the spinal ganglia (Fig. 638) the nerve cells are much larger and for the most part collected in groups near the periphery, while the fibers, which are mostly medullated, traverse the central portion of the ganglion; whereas in the sympathetic ganglia (Fig. 639) the cells are smaller and distributed in irregular groups throughout the whole ganglion; the fibers also are irregularly scattered; some of the entering ones are medullated, while many of those leaving the ganglion are non-medullated. | 31 |
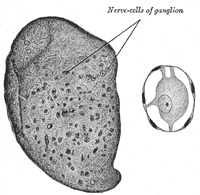 |
FIG. 639– Transverse section of sympathetic ganglion of cat. A. Ganglion. X 50. a. A nerve cell. X 250. (See enlarged image) |
| |
| |
| Neuron Theory.—The nerve cell and its processes collectively constitute what is termed a neuron, and Waldeyer formulated the theory that the nervous system is built up of numerous neurons, “anatomically and genetically independent of one another.” According to this theory (neuron theory) the processes of one neuron only come into contact, and are never in direct continuity, with those of other neurons; while impulses are transmitted from one nerve cell to another through these points of contact, the synapses. The synapse or synaptic membrane seems to allow nervous impulses to pass in one direction only, namely, from the terminals of the axis-cylinder to the dendrons. This theory is based on the following facts, viz.: (1) embryonic nerve cells or neuroblasts are entirely distinct from one another; (2) when nervous tissues are stained by the Golgi method no continuity is seen even between neighboring neurons; and (3) when degenerative changes occur in nervous tissue, either as the result of disease or experiment, they never spread from one neuron to another, but are limited to the individual neurons, or groups of neurons, primarily affected. It must, however, be added that within the past few years the validity of the neuron theory has been called in question by certain eminent histologists, who maintain that by the employment of more delicate histological methods, minute fibrils can be followed from one nerve cell into another. Their existence, however, in the living is open to question. Mott and Marinesco made careful examinations of living cells, using even the ultramicroscope and agree that neither Nissl bodies nor neurofibrils are present in the living state. | 32 |
| For the present we may look upon the neurons as the units or structural elements of the nervous system. All the neurons are present at birth which are present in the adult, their division ceases before birth; they are not all functionally active at birth, but gradually assume functional activity. There is no indication of any regeneration after the destruction of the cell-body of any individual neuron. | 33 |
| Fasciculi, tracts or fiber systems are groups of axons having homologous origin and homologous distribution (as regards their collaterals, subdivisions and terminals) and are often named in accordance with their origin and termination, the name of the nucleus or the location of the cell body from which the axon or fiber arises preceding that of the nucleus or location of its termination. A given topographical area seldom represents a pure tract, as in most cases fibers of different systems are mixed. | 34 |