| Bloodvessels first make their appearance in several scattered vascular areas which are developed simultaneously between the entoderm and the mesoderm of the yolk-sac, i. e., outside the body of the embryo. Here a new type of cell, the angioblast or vasoformative cell, is differentiated from the mesoderm. These cells as they divide form small, dense syncytial masses which soon join with similar masses by means of fine processes to form plexuses. These plexuses increase both by division and growth of its cells and by the addition of new angioblasts which differentiate from the mesoderm. Within these solid plexuses and also within the isolated masses of angioblasts vacuoles appear through liquefaction of the central part of the syncytium into plasma. The lumen of the bloodvessels thus formed is probably intracellular. The flattened cells at the periphery form the endothelium. The nucleated red blood corpuscles develop either from small masses of the original angioblast left attached to the inner wall of the lumen or directly from the flat endothelial cells. In either case the syncytial mass thus formed projects from and is attached to the wall of the vessel. Such a mass is known as a blood island and hemoglobin gradually accumulates within it. Later the cells on the surface round up, giving the mass a mulberry-like appearance. Then the red blood cells break loose and are carried away in the plasma. Such free blood cells continue to divide. The term blood island was originally used for the syncytial masses of angioblasts found in the area vasculosa, but it is probably best to limit the term to the masses within the lumen from which the red blood cells arise as Sabin 88 has done. Blood islands have been seen in the area vasculosa in the omphalomesenteric vein and arteries, and in the dorsal aorta. | 1 |
| The differentiation of angïoblasts from the mesoderm occurs not only in the area vasculosa but within the embryo and probably most of the larger bloodvessels are developed in situ in this manner. This process of the differentiation of angioblasts from the mesoderm probably ceases in different regions of the embryo at different periods and after its cessation new vessels are formed by sprouts from vessels already laid down in the form of capillary plexuses. | 2 |
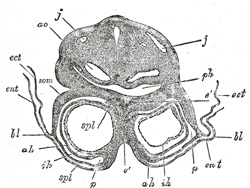 |
FIG. 457– Transverse section through the region of the heart in a rabbit embryo of nine days. X 80. (Kölliker.) j, j. Jugular veins. ao. Aorta. ph. Pharynx. som. Somatopleure. bl. Proamnion. ect. Ectoderm. ent. Entoderm. p. Pericardium. spl. Splanchnopleure. ah. Outer wall of heart. ih. Endothelial lining of heart. é. Septum between heart tubes. (See enlarged image) |
| |
| The first rudiment of the heart appears as a pair of tubular vessels which are developed in the splanchnopleure of the pericardial area (Fig. 457). These are named the primitive aortæ, and a direct continuity is soon established between them and the vessels of the yolk-sac. Each receives anteriorly a vein—the vitelline vein—from the yolk-sac, and is prolonged backward on the lateral aspect of the notochord under the name of the dorsal aorta. The dorsal aortæ give branches to the yolk-sac, and are continued backward through the body-stalk as the umbilical arteries to the villi of the chorion. | 3 |
| Eternod 89 describes the circulation in an embryo which he estimated to be about thirteen days old (Fig. 458). The rudiment of the heart is situated immediately below the fore-gut and consists of a short stem. It gives off two vessels, the primitive aortæ, which run backward, one on either side of the notochord, and then pass into the body-stalk along which they are carried to the chorion. From the chorionic villi the blood is returned by a pair of umbilical veins which unite in the body-stalk to form a single vessel and subsequently encircle the mouth of the yolk-sac and open into the heart. At the junction of the yolk-sac and body-stalk each vein is joined by a branch from the vascular plexus of the yolk-sac. From his observations it seems that, in the human embryo, the chorionic circulation is established before that on the yolk-sac. | 4 |
| By the forward growth and flexure of the head the pericardial area and the anterior portions of the primitive aortæ are folded backward on the ventral aspect of the fore-gut, and the original relation of the somatopleure and splanchnopleure layers of the pericardial area is reversed. Each primitive aorta now consists of a ventral and a dorsal part connected anteriorly by an arch (Fig. 459); these three parts are named respectively the anterior ventral aorta, the dorsal aorta, and the first cephalic arch. The vitelline veins which enter the embryo through the anterior wall of the umbilical orifice are now continuous with the posterior ends of the anterior ventral aorta. With the formation of the tail-fold the posterior parts of the primitive aortæ are carried forward in a ventral direction to form the posterior ventral aortæ and primary caudal arches. 90 In the pericardial region the two primitive aortæ grow together, and fuse to form a single tubular heart (Fig. 460), the posterior end of which receives the two vitelline veins, while from its anterior end the two anterior ventral aortæ emerge. 91 The first cephalic arches pass through the mandibular arches, and behind them five additional pairs subsequently develop, so that altogether six pairs of aortic arches are formed; the fifth arches are very transitory vessels connecting the ventral aortæ with the dorsal ends of the sixth arches. By the rhythmical contraction of the tubular heart the blood is forced through the aortæ and bloodvessels of the vascular area, from which it is returned to the heart by the vitelline veins. This constitutes the vitelline circulation (Fig. 459), and by means of it nutriment is absorbed from the yolk (vitellus.) | 5 |
| The vitelline veins at first open separately into the posterior end of the tubular heart, but after a time their terminal portions fuse to form a single vessel. The vitelline veins ultimately drain the blood from the digestive tube, and are modified to form the portal vein. This is caused by the growth of the liver, which interrupts their direct continuity with the heart; and the blood returned by them circulates through the liver before reaching the heart. | 6 |
 |
FIG. 458– Diagram of the vascular channels in a human embryo of the second week. (After Eternod.) The red lines are the dorsal aortæ continued into the umbilical arteries. The red dotted lines are the ventral aortæ, and the blue dotted lines the vitelline veins. (See enlarged image) |
| |
| With the atrophy of the yolk-sac the vitelline circulation diminishes and ultimately ceases, while an increasing amount of blood is carried through the umbilical arteries to the villi of the chorion. Subsequently, as the non-placental chorionic villi atrophy, their vessels disappear; and then the umbilical arteries convey the whole of their contents to the placenta, whence it is returned to the heart by the umbilical veins. In this manner the placental circulation is established, and by means of it nutritive materials are absorbed from, and waste products given up to the maternal blood. | 7 |
| The umbilical veins, like the vitelline, undergo interruption in the developing liver, and the blood returned by them passes through this organ before reaching the heart. Ultimately the right umbilical vein shrivels up and disappears. | 8 |
| During the occurrence of these changes great alterations take place in the primitive heart and bloodvessels. | 9 |
| |
| Further Development of the Heart.—Between the endothelial lining and the outer wall of the heart there exists for a time an intricate trabecular network of mesodermal tissue from which, at a later stage, the musculi papillares, chordæ tendineæ, and trabeculæ are developed. The simple tubular heart, already described, becomes elongated and bent on itself so as to form an S-shaped loop, the anterior part bending to the right and the posterior part to the left (Fig. 460). The intermediate portion arches transversely from left to right, and then turns sharply forward into the anterior part of the loop. Slight constrictions make their appearance in the tube and divide it from behind forward into five parts. viz.: (1) the sinus venosus; (2) the primitive atrium; (3) the primitive ventricle; (4) the bulbus cordis, and (5) the truncus arteriosus (Figs. 461, 462). The constriction between the atrium and ventricle constitutes the atrial canal, and indicates the site of the future atrioventricular valves. | 10 |
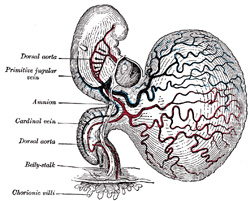 |
FIG. 459– Human embryo of about fourteen days, with yolk-sac. (After His.) (See enlarged image) |
| |
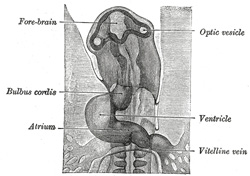 |
FIG. 460– Head of chick embryo of about thirty-eight hours’ incubation, viewed from the ventral surface. X 26 (Duval.) (See enlarged image) |
| |
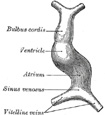 |
FIG. 461– Diagram to illustrate the simple tubular condition of the heart. (Drawn from Ecker-Ziegler model.) (See enlarged image) |
| |
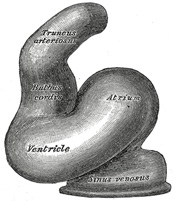 |
FIG. 462– Heart of human embryo of about fourteen days. (From model by His.) (See enlarged image) |
| |
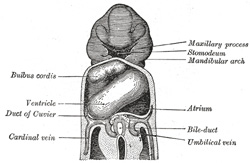 |
FIG. 463– Heart of human embryo of about fifteen days. (Reconstruction by His.) (See enlarged image) |
| |
| The sinus venosus is at first situated in the septum transversum (a layer of mesoderm in which the liver and the central tendon of the diaphragm are developed) behind the primitive atrium, and is formed by the union of the vitelline veins. The veins or ducts of Cuvier from the body of the embryo and the umbilical veins from the placenta subsequently open into it (Fig. 463). The sinus is at first place transversely, and opens by a median aperture into the primitive atrium. Soon, however, it assumes an oblique position, and becomes crescentic in form; its right half or horn increases more rapidly than the left, and the opening into the atrium now communicates with the right portion of the atrial cavity. The right horn and transverse portion of the sinus ultimately become incorporated with and form a part of the adult right atrium, the line of union between it and the auricula being indicated in the interior of the atrium by a vertical crest, the crista terminalis of His. The left horn, which ultimately receives only the left duct of Cuvier, persists as the coronary sinus (Fig. 464). The vitelline and umbilical veins are soon replaced by a single vessel, the inferior vena cava, and the three veins (inferior vena cava and right and left Cuvierian ducts) open into the dorsal aspect of the atrium by a common slit-like aperture (Fig. 465). The upper part of this aperture represents the opening of the permanent superior vena cava, the lower that of the inferior vena cava, and the intermediate part the orifice of the coronary sinus. The slit-like aperture lies obliquely, and is guarded by two halves, the right and left venous valves; above the opening these unite with each other and are continuous with a fold named the septum spurium; below the opening they fuse to form a triangular thickening—the spina vestibuli. The right venous valve is retained; a small septum, the sinus septum, grows from the posterior wall of the sinus venosus to fuse with the valve and divide it into two parts—an upper, the valve of the inferior vena cava, and a lower, the valve of the coronary sinus (Fig. 468). The extreme upper portion of the right venous valve, together with the septum spurium, form the crista terminalis already mentioned. The upper and middle thirds of the left venous valve disappear; the lower third is continued into the spina vestibuli, and later fuses with the septum secundum of the atria and takes part in the formation of the limbus fossæ ovalis. | 11 |
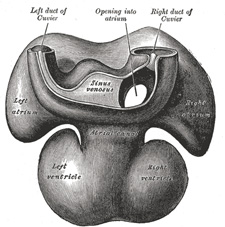 |
FIG. 464– Dorsal surface of heart of human embryo of thirty-five days. (From model by His.) (See enlarged image) |
| |
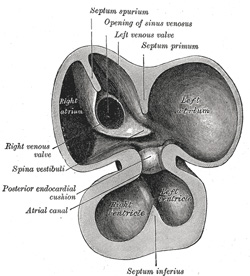 |
FIG. 465– Interior of dorsal half of heart from a human embryo of about thirty days. (From model by His.) (See enlarged image) |
| |
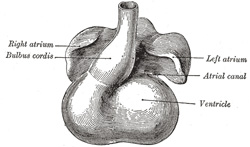 |
FIG. 466– Heart showing expansion of the atria. (Drawn from Ecker-Zeigler model.) (See enlarged image) |
| |
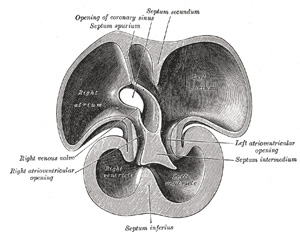 |
FIG. 467– Interior of dorsal half of heart of human embryo of about thirty-five days. (From model by His.) (See enlarged image) |
| |
| The atrial canal is at first a short straight tube connecting the atrial with the ventricular portion of the heart, but its growth is relatively slow, and it becomes overlapped by the atria and ventricles so that its position on the surface of the heart is indicated only by an annular constriction (Fig. 466). Its lumen is reduced to a transverse slit, and two thickenings appear, one on its dorsal and another on its ventral wall. These thickenings, or endocardial cushions (Fig. 465) as they are termed, project into the canal, and, meeting in the middle line, unite to form the septum intermedium which divides the canal into two channels, the future right and left atrioventricular orifices. | 12 |
| The primitive atrium grows rapidly and partially encircles the bulbus cordis; the groove against which the bulbus cordis lies is the first indication of a division into right and left atria. The cavity of the primitive atrium becomes subdivided into right and left chambers by a septum, the septum primum (Fig. 465), which grows downward into the cavity. For a time the atria communicate with each other by an opening, the ostium primum of Born, below the free margin of the septum. This opening is closed by the union of the septum primum with the septum intermedium, and the communication between the atria is reëstablished through an opening which is developed in the upper part of the septum primum; this opening is known as the foramen ovale (ostium secundum of Born) and persists until birth. A second septum, the septum secundum (Figs. 467, 468), semilunar in shape, grows downward from the upper wall of the atrium immediately to the right of the primary septum and foramen ovale. Shortly after birth it fuses with the primary septum, and by this means the foramen ovale is closed, but sometimes the fusion is incomplete and the upper part of the foramen remains patent. The limbus fossæ ovalis denotes the free margin of the septum secundum. Issuing from each lung is a pair of pulmonary veins; each pair unites to form a single vessel, and these in turn join in a common trunk which opens into the left atrium. Subsequently the common trunk and the two vessels forming it expand and form the vestibule or greater part of the atrium, the expansion reaching as far as the openings of the four vessels, so that in the adult all four veins open separately into the left atrium. | 13 |
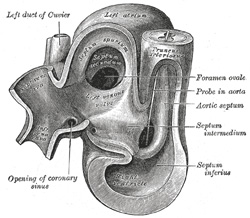 |
FIG. 468– Same heart as in Fig. 467, opened on right side. (From model by His.) (See enlarged image) |
| |
| The primitive ventricle becomes divided by a septum, the septum inferius or ventricular septum (Figs. 465, 466, 467), which grows upward from the lower part of the ventricle, its position being indicated on the surface of the heart by a furrow. Its dorsal part increases more rapidly than its ventral portion, and fuses with the dorsal part of the septum intermedium. For a time an interventricular foramen exists above its ventral portion (Fig. 468), but this foramen is ultimately closed by the fusion of the aortic septum with the ventricular septum. | 14 |
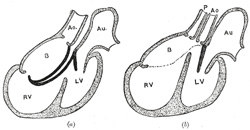 |
FIG. 469– Diagrams to illustrate the transformation of the bulbus cordis. (Keith.) Ao. Truncus arteriosus. Au. Atrium. B. Bulbus cordis. RV. Right ventricle. LV. Left ventricle. P. Pulmonary artery. (See enlarged image) |
| |
| When the heart assumes its S-shaped form the bulbus cordis lies ventral to and in front of the primitive ventricle. The adjacent walls of the bulbus cordis and ventricle approximate, fuse, and finally disappear, and the bulbus cordis now communicates freely with the right ventricle, while the junction of the bulbus with the truncus arteriosus is brought directly ventral to and applied to the atrial canal. By the upgrowth of the ventricular septum the bulbus cordis is in great measure separated from the left ventricle, but remains an integral part of the right ventricle, of which it forms the infundibulum (Fig. 469). | 15 |
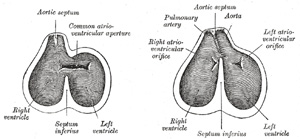 |
FIG. 470– Diagrams to show the development of the septum of the aortic bulb and of the ventricles. (Born.) (See enlarged image) |
| |
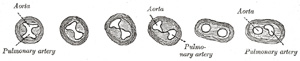 |
FIG. 471– Transverse sections through the aortic bulb to show the growth of the aortic septum. The lowest section is on the left, the highest on the right of the figure. (After His.) (See enlarged image) |
| |
| The truncus arteriosus and bulbus cordis are divided by the aortic septum (Fig. 470). This makes its appearance in three portions. (1) Two distal ridge-like thickenings project into the lumen of the tube; these increase in size, and ultimately meet and fuse to form a septum, which takes a spiral course toward the proximal end of the truncus arteriosus. It divides the distal part of the truncus into two vessels, the aorta and pulmonary artery, which lie side by side above, but near the heart the pulmonary artery is in front of the aorta. (2) Four endocardial cushions appear in the proximal part of the truncus arteriosus in the region of the future semilunar valves; the manner in which these are related to the aortic septum is described below. (3) Two endocardial thickenings—anterior and posterior—develop in the bulbus cordis and unite to form a short septum; this joins above with the aortic septum and below with the ventricular septum. The septum grows down into the ventricle as an oblique partition, which ultimately blends with the ventricular septum in such a way as to bring the bulbus cordis into communication with the pulmonary artery, and through the latter with the sixth pair of aortic arches; while the left ventricle is brought into continuity with the aorta, which communicates with the remaining aortic arches. | 16 |
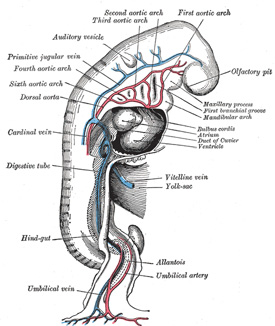 |
FIG. 472– Profile view of a human embryo estimated at twenty or twenty-one days old. (After His.) (See enlarged image) |
| |
| |
| The Valves of the Heart.—The atrioventricular valves are developed in relation to the atrial canal. By the upward expansion of the bases of the ventricles the canal becomes invaginated into the ventricular cavities. The invaginated margin forms the rudiments of the lateral cusps of the atrioventricular valves; the mesial or septal cusps of the valves are developed as downward prolongations of the septum intermedium (Fig. 467). The aortic and pulmonary semilunar valves are formed from four endocardial thickenings—an anterior, a posterior, and two lateral—which appear at the proximal end of the truncus arteriosus. As the aortic septum grows downward it divides each of the lateral thickenings into two, thus giving rise to six thickenings—the rudiments of the semilunar valves—three at the aortic and three at the pulmonary orifice (Fig. 471). | 17 |
| |
| Further Development of the Arteries.—Recent observations show that practically none of the main vessels of the adult arise as such in the embryo. In the site of each vessel a capillary network forms, and by the enlargement of definite paths in this the larger arteries and veins are developed. The branches of the main arteries are not always simple modifications of the vessels of the capillary network, but may arise as new outgrowths from the enlarged stem. | 18 |
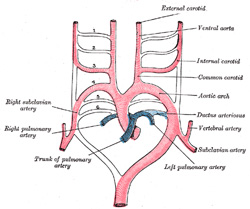 |
FIG. 473– Scheme of the aortic arches and their destination. (Modified from Kollmann.) (See enlarged image) |
| |
| It has been seen (page 506) that each primitive aorta consists of a ventral and a dorsal part which are continuous through the first aortic arch. The dorsal aortæ at first run backward separately on either side of the notochord, but about the third week they fuse from about the level of the fourth thoracic to that of the fourth lumbar segment to form a single trunk, the descending aorta. The first aortic arches run through the mandibular arches, and behind them five additional pairs are developed within the visceral arches; so that, in all, six pairs of aortic arches are formed (Figs. 472, 473). The first and second arches pass between the ventral and dorsal aortæ, while the others arise at first by a common trunk from the truncus arteriosus, but end separately in the dorsal aortæ. As the neck elongates, the ventral aortæ are drawn out, and the third and fourth arches arise directly from these vessels. | 19 |
| In fishes these arches persist and give off branches to the gills, in which the blood is oxygenated. In mammals some of them remain as permanent structures while others disappear or become obliterated (Fig. 473). | 20 |
| |
| The Anterior Ventral Aortæ.—These persist on both sides. The right forms (a) the innominate artery, (b) the right common and external carotid arteries. The left gives rise to (a) the short portion of the aortic arch, which reaches from the origin of the innominate artery to that of the left common carotid artery; (b) the left common and external carotid arteries. | 21 |
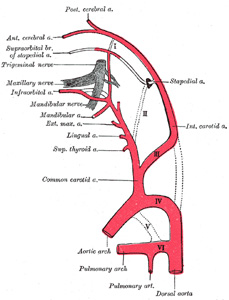 |
FIG. 474– Diagram showing the origins of the main branches of the carotid arteries. (Founded on Tandler.) (See enlarged image) |
| |
| |
| The Aortic Arches.—The first and second arches disappear early, but the dorsal end of the second gives origin to the stapedial artery (Fig. 474), a vessel which atrophies in man but persists in some mammals. It passes through the ring of the stapes and divides into supraorbital, infraorbital, and mandibular branches which follow the three divisions of the trigeminal nerve. The infraorbital and mandibular arise from a common stem, the terminal part of which anastomoses with the external carotid. On the obliteration of the stapedial artery this anastomosis enlarges and forms the internal maxillary artery, and the branches of the stapedial artery are now branches of this vessel. The common stem of the infraorbital and mandibular branches passes between the two roots of the auriculotemporal nerve and becomes the middle meningeal artery; the original supraorbital branch of the stapedial is represented by the orbital twigs of the middle meningeal. The third aortic arch constitutes the commencement of the internal carotid artery, and is therefore named the carotid arch. The fourth right arch forms the right subclavian as far as the origin of its internal mammary branch; while the fourth left arch constitutes the arch of the aorta between the origin of the left carotid artery and the termination of the ductus arteriosus. The fifth arch disappears on both sides. The sixth right arch disappears; the sixth left arch gives off the pulmonary arteries and forms the ductus arteriosus; this duct remains pervious during the whole of fetal life, but is obliterated a few days after birth. His showed that in the early embryo the right and left arches each gives a branch to the lungs, but that later both pulmonary arteries take origin from the left arch. | 22 |
| |
| The Dorsal Aortæ.—In front of the third aortic arches the dorsal aortæ persist and form the continuations of the internal carotid arteries; these arteries pass to the brain and each divides into an anterior and a posterior branch, the former giving off the ophthalmic and the anterior and middle cerebral arteries, while the latter turns back and joins the cerebral part of the vertebral artery. Behind the third arch the right dorsal aorta disappears as far as the point where the two dorsal aortæ fuse to form the descending aorta. The part of the left dorsal aorta between the third and fourth arches disappears, while the remainder persists to form the descending part of the arch of the aorta. A constriction, the aortic isthmus, is sometimes seen in the aorta between the origin of the left subclavian and the attachment of the ductus arteriosus. | 23 |
| Sometimes the right subclavian artery arises from the aortic arch distal to the origin of the left subclavian and passes upward and to the right behind the trachea and esophagus. This condition may be explained by the persistence of the right dorsal aorta and the obliteration of the fourth right arch. | 24 |
| In birds the fourth right arch forms the arch of the aorta; in reptiles the fourth arch on both sides persists and gives rise to the double aortic arch in these animals. | 25 |
| The heart originally lies on the ventral aspect of the pharynx, immediately behind the stomodeum. With the elongation of the neck and the development of the lungs it recedes within the thorax, and, as a consequence, the anterior ventral aortæ are drawn out and the original position of the fourth and fifth arches is greatly modified. Thus, on the right side the fourth recedes to the root of the neck, while on the left side it is withdrawn within the thorax. The recurrent nerves originally pass to the larynx under the sixth pair of arches, and are therefore pulled backward with the descent of these structures, so that in the adult the left nerve hooks around the ligamentum arteriosum; owing to the disappearance of the fifth and the sixth right arches the right nerve hooks around that immediately above them, i.e., the commencement of the subclavian artery. Segmental arteries arise from the primitive dorsal aortæ and course between successive segments. The seventh segmental artery is of special interest, since it forms the lower end of the vertebral artery and, when the forelimb bud appears, sends a branch to it (the subclavian artery). From the seventh segmental arteries the entire left subclavian and the greater part of the right subclavian are formed. The second pair of segmental arteries accompany the hypoglossal nerves to the brain and are named the hypoglossal arteries. Each sends forward a branch which forms the cerebral part of the vertebral artery and anastomoses with the posterior branch of the internal carotid. The two vertebrals unite on the ventral surface of the hind-brain to form the basilar artery. Later the hypoglossal artery atrophies and the vertebral is connected with the first segmental artery. The cervical part of the vertebral is developed from a longitudinal anastomosis between the first seven segmental arteries, so that the seventh of these ultimately becomes the source of the artery. As a result of the growth of the upper limb the subclavian artery increases greatly in size and the vertebral then appears to spring from it. | 26 |
| Recent observations show that several segmental arteries contribute branches to the upper limb-bud and form in it a free capillary anastomosis. Of these branches, only one, viz., that derived from the seventh segmental artery, persists to form the subclavian artery. The subclavian artery is prolonged into the limb under the names of the axillary and brachial arteries, and these together constitute the arterial stem for the upper arm, the direct continuation of this stem in the forearm is the volar interosseous artery. A branch which accompanies the median nerve soon increases in size and forms the main vessel (median artery) of the forearm, while the volar interosseous diminishes. Later the radial and ulnar arteries are developed as branches of the brachial part of the stem and coincidently with their enlargement the median artery recedes; occasionally it persists as a vessel of some considerable size and then accompanies the median nerve into the palm of the hand. | 27 |
| The primary arterial stem for the lower limb is formed by the inferior gluteal (sciatic) artery, which accompanies the sciatic nerve along the posterior aspect of the thigh to the back of the knee, whence it is continued as the peroneal artery. This arrangement exists in reptiles and amphibians. The femoral artery arises later as a branch of the common iliac, and, passing down the front and medial side of the thigh to the bend of the knee, joins the inferior gluteal artery. The femoral quickly enlarges, and, coincidently with this, the part of the inferior gluteal immediately above the knee undergoes atrophy. The anterior and posterior tibial arteries are branches of the main arterial stem. | 28 |
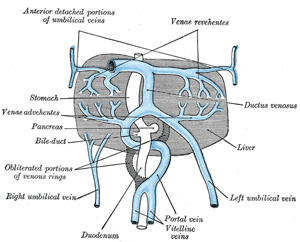 |
FIG. 475– The liver and the veins in connection with it, of a human embryo, twenty-four or twenty-five days old, as seen from the ventral surface. (After His.) (See enlarged image) |
| |
| |
| Further Development of the Veins.—The formation of the great veins of the embryo may be best considered by dividing them into two groups, visceral and parietal. | 29 |
| |
| The Visceral Veins.—The visceral veins are the two vitelline or omphalomesenteric veins bringing the blood from the yolk-sac, and the two umbilical veins returning the blood from the placenta; these four veins open close together into the sinus venosus. | 30 |
| The Vitelline Veins run upward at first in front, and subsequently on either side of the intestinal canal. They unite on the ventral aspect of the canal, and beyond this are connected to one another by two anastomotic branches, one on the dorsal, and the other on the ventral aspect of the duodenal portion of the intestine, which is thus encircled by two venous rings (Fig. 475); into the middle or dorsal anastomosis the superior mesenteric vein opens. The portions of the veins above the upper ring become interrupted by the developing liver and broken up by it into a plexus of small capillary-like vessels termed sinusoids (Minot). The branches conveying the blood to this plexus are named the venæ advehentes, and become the branches of the portal vein; while the vessels draining the plexus into the sinus venosus are termed the venæ revehentes, and form the future hepatic veins (Figs. 475, 476). Ultimately the left vena revehens no longer communicates directly with the sinus venosus, but opens into the right vena revehens. The persistent part of the upper venous ring, above the opening of the superior mesenteric vein, forms the trunk of the portal vein. | 31 |
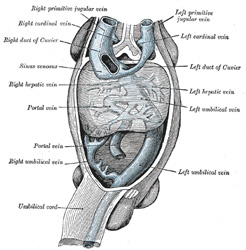 |
FIG. 476– Human embryo with heart and anterior body-wall removed to show the sinus venosus and its tributaries. (After His.) (See enlarged image) |
| |
| The two Umbilical Veins fuse early to form a single trunk in the body-stalk, but remain separate within the embryo and pass forward to the sinus venosus in the side walls of the body. Like the vitelline veins, their direct connection with the sinus venosus becomes interrupted by the developing liver, and thus at this stage the whole of the blood from the yolk-sac and placenta passes through the substance of the liver before it reaches the heart. The right umbilical and right vitelline veins shrivel and disappear; the left umbilical, on the other hand, becomes enlarged and opens into the upper venous ring of the vitelline veins; with the atrophy of the yolk-sac the left vitelline vein also undergoes atrophy and disappears. Finally a direct branch is established between this ring and the right hepatic vein; this branch is named the ductus venosus, and, enlarging rapidly, it forms a wide channel through which most of the blood, returned from the placenta, is carried direct to the heart without passing through the liver. A small proportion of the blood from the placenta is, however, conveyed from the left umbilical vein to the liver through the left vena advehens. The left umbilical vein and the ductus venosus undergo atrophy and obliteration after birth, and form respectively the ligamentum teres and ligamentum venosum of the liver. | 32 |
| |
| The Parietal Veins.—The first indication of a parietal system consists in the appearance of two short transverse veins, the ducts of Cuvier, which open, one on either side, into the sinus venosus. Each of these ducts receives an ascending and descending vein. The ascending veins return the blood from the parietes of the trunk and from the Wolffian bodies, and are called cardinal veins. The descending veins return the blood from the head, and are called primitive jugular veins (Fig. 477). The blood from the lower limbs is collected by the right and left iliac and hypogastric veins, which, in the earlier stages of development, open into the corresponding right and left cardinal veins; later, a transverse branch (the left common iliac vein) is developed between the lower parts of the two cardinal veins (Fig. 479), and through this the blood is carried into the right cardinal vein. The portion of the left cardinal vein below the left renal vein atrophies and disappears up to the point of entrance of the left spermatic vein; the portion above the left renal vein persists as the hemiazygos and accessory hemiazygos veins and the lower portion of the highest left intercostal vein. The right cardinal vein which now receives the blood from both lower extremities, forms a large venous trunk along the posterior abdominal wall; up to the level of the renal veins it forms the lower part of the inferior vena cava. Above the level of the renal veins the right cardinal vein persists as the azygos vein and receives the right intercostal veins, while the hemiazygos veins are brought into communication with it by the development of transverse branches in front of the vertebral column (Figs. 479, 480) | 33 |
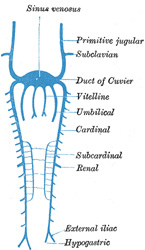 |
FIG. 477– Scheme of arrangement of parietal veins. (See enlarged image) |
| |
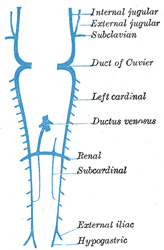 |
FIG. 478– Scheme showing early stages of development of the inferior vena cava. (See enlarged image) |
| |
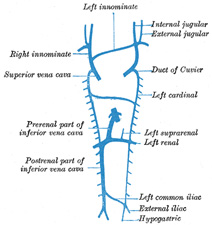 |
FIG. 479– Diagram showing development of main cross branches between jugulars and between cardinals. (See enlarged image) |
| |
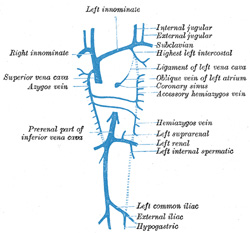 |
FIG. 480– Diagram showing completion of development of the parietal veins. (See enlarged image) |
| |
| |
| Inferior Vena Cava.—The development of the inferior vena cava is associated with the formation of two veins, the subcardinal veins (Figs. 477, 478). These lie parallel to, and on the ventral aspect of, the cardinal veins, and originate as longitudinal anastomosing channels which link up the tributaries from the mesentery to the cardinal veins; they communicate with the cardinal veins above and below, and also by a series of transverse branches. The two subcardinals are for a time connected with each other in front of the aorta by cross branches, but these disappear and are replaced by a single transverse channel at the level where the renal veins join the cardinals, and at the same level a cross communication is established on either side between the cardinal and subcardinal (Fig. 478). The portion of the right subcardinal behind this cross communication disappears, while that in front, i.e., the prerenal part, forms a connection with the ductus venosus at the point of opening of the hepatic veins, and, rapidly enlarging, receives the blood from the postrenal part of the right cardinal through the cross communication referred to. In this manner a single trunk, the inferior vena cava (Fig. 480), is formed, and consists of the proximal part of the ductus venosus, the prerenal part of the right subcardinal vein, the postrenal part of the right cardinal vein, and the cross branch which joins these two veins. The left subcardinal disappears, except the part immediately in front of the renal vein, which is retained as the left suprarenal vein. The spermatic (or ovarian) vein opens into the postrenal part of the corresponding cardinal vein. This portion of the right cardinal, as already explained, forms the lower part of the inferior vena cava, so that the right spermatic opens directly into that vessel. The postrenal segment of the left cardinal disappears, with the exception of the portion between the spermatic and renal vein, which is retained as the terminal part of the left spermatic vein. | 34 |
| In consequence of the atrophy of the Wolffian bodies the cardinal veins diminish in size; the primitive jugular veins, on the other hand, become enlarged, owing to the rapid development of the head and brain. They are further augmented by receiving the veins (subclavian) from the upper extremities, and so come to form the chief veins of the Cuvierian ducts; these ducts gradually assume an almost vertical position in consequence of the descent of the heart into the thorax. The right and left Cuvierian ducts are originally of the same diameter, and are frequently termed the right and left superior venæ cavæ. By the development of a transverse branch, the left innominate vein between the two primitive jugular veins, the blood is carried across from the left to the right primitive jugular (Figs. 479, 480). The portion of the right primitive jugular vein between the left innominate and the azygos vein forms the upper part of the superior vena cava of the adult; the lower part of this vessel, i.e., below the entrance of the azygos vein, is formed by the right Cuvierian duct. Below the origin of the transverse branch the left primitive jugular vein and left Cuvierian duct atrophy, the former constituting the upper part of the highest left intercostal vein, while the latter is represented by the ligament of the left vena cava, vestigial fold of Marshall, and the oblique vein of the left atrium, oblique vein of Marshall (Fig. 480). Both right and left superior venæ cavæ are present in some animals, and are occasionally found in the adult human being. The oblique vein of the left atrium passes downward across the back of the left atrium to open into the coronary sinus, which, as already indicated, represents the persistent left horn of the sinus venosus. | 35 |
| |
| Venous Sinuses of the Dura Mater. 92—The primary arrangement for drainage of the capillaries of the head (Figs. 481, 488) consists of a primary head vein which starts in the region of the midbrain and runs caudalward along the side of the brain tube to terminate at the duct of Cuvier. The primary head vein drains three plexuses of capillaries: the anterior dural plexus, the middle dural plexus and the posterior dural plexus. The growth of the cartilaginous capsule of the ear and the growth and alteration in form of the brain bring about changes in this primary arrangement (Figs. 483–488). Owing to the growth of the otic capsule and middle ear the course of the primary head vein becomes unfavorable and a segment of it becomes obliterated. To make the necessary adjustment an anastomosis is established above the otic capsule (Fig. 483) and the middle plexus drains into the posterior plexus. Then the anteror plexus fuses with the middle plexus (Fig. 484) and drains through it and the newly established channel, dorsal to the otic capsule. All that remains of the primary head vein is the cardinal portion or internal jugular and the part in the region of the trigeminal nerve which may be called the cavernous sinus. Into it drain the orbital veins. The drainage from the cavernous sinus is now upward through the original trunk of the middle plexus, which is now the superior petrosal sinus, into the newly established dorsal channel. This dorsal channel is the transverse sinus (Figs. 485–488). The inferior petrosal sinus appears later (Fig. 486). From the anterior plexus a sagittal plexus extends forward from which develops the superior sagittal sinus (Figs. 484–488). The straight sinus is formed in the ventral part of the sagittal plexus. As the hemispheres extend backward these sinuses elongate by incorporating the more caudal loops of the plexus. The anterior part of the sinus is completed first. | 36 |
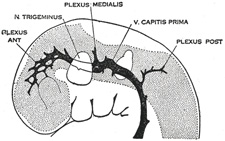 |
FIG. 481– 4 mm. (See enlarged image) |
| |
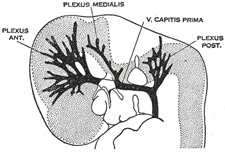 |
FIG. 482– 14 mm. (See enlarged image) |
| |
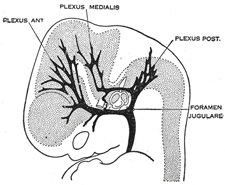 |
FIG. 483– 18 mm. (See enlarged image) |
| |
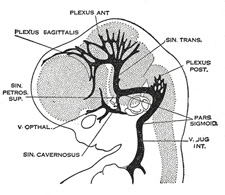 |
FIG. 484– 21 mm. (See enlarged image) |
| |
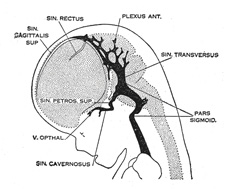 |
FIG. 485– 35 mm. (See enlarged image) |
| |
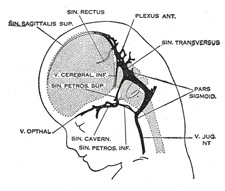 |
FIG. 486– 50 mm. crown-rump length. (See enlarged image) |
| |
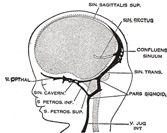 |
FIG. 487– 80 mm. crown-rump length.(After Streeter.) (See enlarged image) |
| |
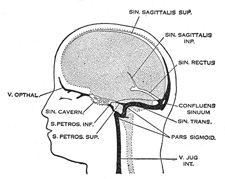 |
FIG. 488– Profile drawings of the dural veins showing principal stages in their development in human embryos from 4 mm. to birth. It is of particular interest to notice their adaptation to the growth and changes in the form of the central nervous system. Fig. 481, 4 mm.; Fig. 482, 14 mm.; Fig. 483, 18 mm.; Fig. 484, 21 mm.; Fig. 485, 35 mm.; Fig. 486, 50 mm. crown-rump length; Fig. 487, 80 mm. crown-rump length; Fig. 488, adult. (After Streeter.) (See enlarged image) |
| |
| The external jugular vein at first drains the region behind the ear (posterior auricular) and enters the primitive jugular as a lateral tributary. A group of veins from the face and lingual region converge to form a common vein, the linguo-facial, 93 which also terminates in the primitive jugular. Later, cross communications develop between the external jugular and the linguo-facial, with the result that the posterior group of facial veins is transferred to the external jugular. | 37 |