| The hind-brain or rhombencephalon occupies the posterior fossa of the cranial cavity and lies below a fold of dura mater, the tentorium cerebelli. It consists of (a) the myelencephalon, comprising the medulla oblongata and the lower part of the fourth ventricle; (b) the metencephalon, consisting of the pons, cerebellum, and the intermediate part of the fourth ventricle; and (c) the isthmus rhombencephali, a constricted portion immediately adjoining the mid-brain and including the superior peduncles of the cerebellum, the anterior medullary velum, and the upper part of the fourth ventricle. | 1 |
| |
| The Medulla Oblongata (spinal bulb).—The medulla oblongata extends from the lower margin of the pons to a plane passing transversely below the pyramidal decussation and above the first pair of cervical nerves; this plane corresponds with the upper border of the atlas behind, and the middle of the odontoid process of the axis in front; at this level the medulla oblongata is continuous with the medulla spinalis. Its anterior surface is separated from the basilar part of the occipital bone and the upper part of the odontoid process by the membranes of the brain and the occipitoaxial ligaments. Its posterior surface is received into the fossa between the hemispheres of the cerebellum, and the upper portion of it forms the lower part of the floor of the fourth ventricle. | 2 |
| The medulla oblongata is pyramidal in shape, its broad extremity being directed upward toward the pons, while its narrow, lower end is continuous with the medulla spinalis. It measures about 3 cm. in length, about 2 cm. in breadth at its widest part, and about 1.25 cm. in thickness. The central canal of the medulla spinalis is prolonged into its lower half, and then opens into the cavity of the fourth ventricle; the medulla oblongata may therefore be divided into a lower closed part containing the central canal, and an upper open part corresponding with the lower portion of the fourth ventricle. | 3 |
| The Anterior Median Fissure (fissura mediana anterior; ventral or ventromedian fissure) contains a fold of pia mater, and extends along the entire length of the medulla oblongata: it ends at the lower border of the pons in a small triangular expansion, termed the foramen cecum. Its lower part is interrupted by bundles of fibers which cross obliquely from one side to the other, and constitute the pyramidal decussation. Some fibers, termed the anterior external arcuate fibers, emerge from the fissure above this decussation and curve lateralward and upward over the surface of the medulla oblongata to join the inferior peduncle. | 4 |
| The Posterior Median Fissure (fissura mediana posterior; dorsal or dorsomedian fissure) is a narrow groove; and exists only in the closed part of the medulla oblongata; it becomes gradually shallower from below upward, and finally ends about the middle of the medulla oblongata, where the central canal expands into the cavity of the fourth ventricle. | 5 |
| These two fissures divide the closed part of the medulla oblongata into symmetrical halves, each presenting elongated eminences which, on surface view, are continuous with the funiculi of the medulla spinalis. In the open part the halves are separated by the anterior median fissure, and by a median raphé which extends from the bottom of the fissure to the floor of the fourth ventricle. Further, certain of the cranial nerves pass through the substance of the medulla oblongata, and are attached to its surface in series with the roots of the spinal nerves; thus, the fibers of the hypoglossal nerve represent the upward continuation of the anterior nerve roots, and emerge in linear series from a furrow termed the antero-lateral sulcus. Similarly, the accessory, vagus, and glossopharyngeal nerves correspond with the posterior nerve roots, and are attached to the bottom of a sulcus named the postero-lateral sulcus. Advantage is taken of this arrangement to subdivide each half of the medulla oblongata into three districts, anterior, middle, and posterior. Although these three districts appear to be directly continuous with the corresponding funiculi of the medulla spinalis, they do not necessarily contain the same fibers, since some of the fasciculi of the medulla spinalis end in the medulla oblongata, while others alter their course in passing through it. | 6 |
| The anterior district (Fig. 679) is named the pyramid (pyramis medullæ oblongatæ) and lies between the anterior median fissure and the antero-lateral sulcus. Its upper end is slightly constricted, and between it and the pons the fibers of the abducent nerve emerge; a little below the pons it becomes enlarged and prominent, and finally tapers into the anterior funiculus of the medulla spinalis, with which, at first sight, it appears to be directly continuous. | 7 |
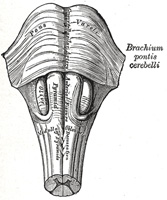 |
FIG. 679– Medulla oblongata and pons. Anterior surface. (See enlarged image) |
| |
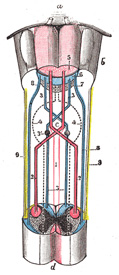 |
FIG. 680– Decussation of pyramids. Scheme showing passage of various fasciculi from medulla spinalis to medulla oblongata. a. Pons. b. Medulla oblongata. c. Decussation of the pyramids. d. Section of cervical part of medulla spinalis. 1. Anterior cerebrospinal fasciculus (in red). 2. Lateral cerebrospinal fasciculus (in red). 3. Sensory tract (fasciculi gracilis et cuneatus) (in blue). 3’. Gracile and cuneate nuclei. 4. Antero-lateral proper fasciculus (in dotted line). 5. Pyramid. 6. Lemniscus. 7. Medial longitudinal fasciculus. 8. Ventral spinocerebellar fasciculus (in blue). 9. Dorsal spinocerebellar fasciculus (in yellow). (Testut.) (See enlarged image) |
| |
| The two pyramids contain the motor fibers which pass from the brain to the medulla oblongata and medulla spinalis, corticobulbar and corticospinal fibers. When these pyramidal fibers are traced downward it is found that some two-thirds or more of them leave the pyramids in successive bundles, and decussate in the anterior median fissure, forming what is termed the pyramidal decussation. Having crossed the middle line, they pass down in the posterior part of the lateral funiculus as the lateral cerebrospinal fasciculus. The remaining fibers—i. e., those which occupy the lateral part of the pyramid—do not cross the middle line, but are carried downward as the anterior cerebrospinal fasciculus (Fig. 680) into the anterior funiculus of the same side. | 8 |
| The greater part of the anterior proper fasciculus of the medulla spinalis is continued upward through the medulla oblongata under the name of the medial longitudinal fasciculus. | 9 |
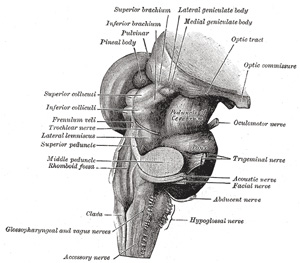 |
FIG. 681– Hind- and mid-brains; postero-lateral view. (See enlarged image) |
| |
| The lateral district (Fig. 681) is limited in front by the antero-lateral sulcus and the roots of the hypoglossal nerve, and behind by the postero-lateral sulcus and the roots of the accessory, vagus, and glossopharyngeal nerves. Its upper part consists of a prominent oval mass which is named the olive, while its lower part is of the same width as the lateral funiculus of the medulla spinalis, and appears on the surface to be a direct continuation of it. As a matter of fact, only a portion of the lateral funiculus is continued upward into this district, for the lateral cerebrospinal fasciculus passes into the pyramid of the opposite side, and the dorsal spinocerebellar fasciculus is carried into the inferior peduncle in the posterior district. The ventral spinocerebellar fasciculus is continued upward on the lateral surface of the medulla oblongata in the same relative position it occupies in the spinal cord until it passes under cover of the external arcuate fibers. It passes beneath these fibers just dorsal to the olive and ventral to the roots of the vagus and glossopharyngeal nerves; it continues upward through the pons along the dorso-lateral edge of the lateral lemniscus. The remainder of the lateral funiculus consists chiefly of the lateral proper fasciculus. Most of these fibers dip beneath the olive and disappear from the surface; but a small strand remains superficial to the olive. In a depression at the upper end of this strand is the acoustic nerve. | 10 |
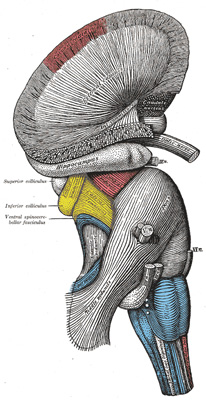 |
FIG. 682– Superficial dissection of brain-stem. Lateral view. (See enlarged image) |
| |
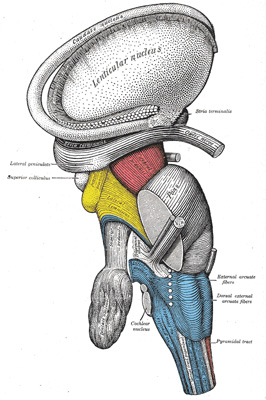 |
FIG. 683– Dissection of brain-stem. Lateral view. (See enlarged image) |
| |
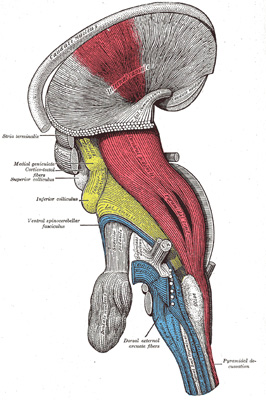 |
FIG. 684– Deep dissection of brain-stem. Lateral view. (See enlarged image) |
| |
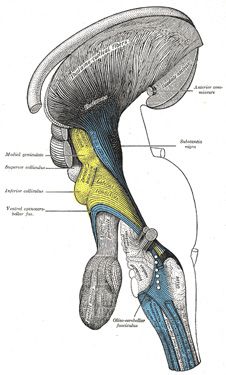 |
FIG. 685– Deep dissection of brain-stem. Lateral view. (See enlarged image) |
| |
| The olive (oliva; olivary body) is situated lateral to the pyramid, from which it is separated by the antero-lateral sulcus, and the fibers of the hypoglossal nerve. Behind, it is separated from the postero-lateral sulcus by the ventral spinocerebellar fasciculus. In the depression between the upper end of the olive and the pons lies the acoustic nerve. It measures about 1.25 cm. in length, and between its upper end and the pons there is a slight depression to which the roots of the facial nerve are attached. The external arcuate fibers wind across the lower part of the pyramid and olive and enter the inferior peduncle. | 11 |
| The posterior district (Fig. 686) lies behind the postero-lateral sulcus and the roots of the accessory, vagus, and the glossopharyngeal nerves, and, like the lateral district, is divisible into a lower and an upper portion. | 12 |
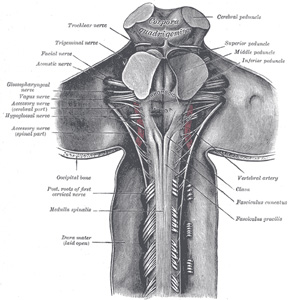 |
FIG. 686– Upper part of medulla spinalis and hind- and mid-brains; posterior aspect, exposed in situ. (See enlarged image) |
| |
| The lower part is limited behind by the posterior median fissure, and consists of the fasciculus gracilis and the fasciculus cuneatus. The fasciculus gracilis is placed parallel to and along the side of the posterior median fissure, and separated from the fasciculus cuneatus by the postero-intermediate sulcus and septum. The gracile and cuneate fasciculi are at first vertical in direction; but at the lower part of the rhomboid fossa they diverge from the middle line in a V-shaped manner, and each presents an elongated swelling. That on the fasciculus gracilis is named the clava, and is produced by a subjacent nucleus of gray matter, the nucleus gracilis; that on the fasciculus cuneatus is termed the cuneate tubercle, and is likewise caused by a gray nucleus, named the nucleus cuneatus. The fibers of these fasciculi terminate by arborizing around the cells in their respective nuclei. A third elevation, produced by the substantia gelatinosa of Rolando, is present in the lower part of the posterior district of the medulla oblongata. It lies on the lateral aspect of the fasciculus cuneatus, and is separated from the surface of the medulla oblongata by a band of nerve fibers which form the spinal tract (spinal root) of the trigeminal nerve. Narrow below, this elevation gradually expands above, and ends, about 1.25 cm. below the pons, in a tubercle, the tubercle of Rolando (tuber cinereum). | 13 |
| The upper part of the posterior district of the medulla oblongata is occupied by the inferior peduncle, a thick rope-like strand situated between the lower part of the fourth ventricle and the roots of the glossopharyngeal and vagus nerves. The inferior peduncles connect the medulla spinalis and medulla oblongata with the cerebellum, and are sometimes named the restiform bodies. As they pass upward, they diverge from each other, and assist in forming the lower part of the lateral boundaries of the fourth ventricle; higher up, they are directed backward, each passing to the corresponding cerebellar hemisphere. Near their entrance, into the cerebellum they are crossed by several strands of fibers, which run to the median sulcus of the rhomboid fossa, and are named the striæ medullares. The inferior peduncle appears to be the upward continuation of the fasciculus gracilis and fasciculus cuneatus; this, however, is not so, as the fibers of these fasciculi end in the gracile and cuneate nuclei. The constitution of the inferior peduncle will be subsequently discussed. | 14 |
| Caudal to the striæ medullares the inferior peduncle is partly covered by the corpus pontobulbare (Essick 120), a thin mass of cells and fibers extending from the pons between the origin of the VII and VIII cranial nerves. | 15 |
| |
| Internal Structure of the Medulla Oblongata.—Although the external form of the medulla oblongata bears a certain resemblance to that of the upper part of the medulla spinalis, its internal structure differs widely from that of the latter, and this for the following principal reasons: (1) certain fasciculi which extend from the medulla spinalis to the brain, and vice versa, undergo a rearrangement in their passage through the medulla oblongata; (2) others which exist in the medulla spinalis end in the medulla oblongata; (3) new fasciculi originate in the gray substance of the medulla oblongata and pass to different parts of the brain; (4) the gray substance, which in the medulla spinalis forms a continuous H-shaped column, becomes greatly modified and subdivided in the medulla oblongata, where also new masses of gray substance are added; (5) on account of the opening out of the central canal of the medulla spinalis, certain parts of the gray substance, which in the medulla spinalis were more or less centrally situated, are displayed in the rhomboid fossa; (6) the medulla oblongata is intimately associated with many of the cranial nerves, some arising form, and others ending in, nuclei within its substance. | 16 |
| |
| The Cerebrospinal Fasciculi.—The downward course of these fasciculi from the pyramids of the medulla oblongata and their partial decussation have already been described (page 761). In crossing to reach the lateral funiculus of the opposite side, the fibers of the lateral cerebrospinal fasciculi extend backward through the anterior columns, and separate the head of each of these columns from its base (Figs. 687, 688). The base retains its position in relation to the ventral aspect of the central canal, and, when the latter opens into the fourth ventricle, appears in the rhomboid fossa close to the middle line, where it forms the nuclei of the hypoglossal and abducent nerves; while above the level of the ventricle it exists as the nuclei of the trochlear and oculomotor nerves in relation to the floor of the cerebral aqueduct. The head of the column is pushed lateralward and forms the nucleus ambiguus, which gives origin from below upward to the cranial part of the accessory and the motor fibers of the vagus and glossopharyngeal, and still higher to the motor fibers of the facial and trigeminal nerves. | 17 |
| The fasciculus gracilis and fasciculus cuneatus constitute the posterior sensory fasciculi of the medulla spinalis; they are prolonged upward into the lower part of the medulla oblongata, where they end respectively in the nucleus gracilis and nucleus cuneatus. These two nuclei are continuous with the central gray substance of the medulla spinalis, and may be regarded as dorsal projections of this, each being covered superficially by the fibers of the corresponding fasciculus. On transverse section (Fig. 694) the nucleus gracilis appears as a single, more or less quadrangular mass, while the nucleus cuneatus consists of two parts: a larger, somewhat triangular, medial nucleus, composed of small or medium-sized cells, and a smaller lateral nucleus containing large cells. | 18 |
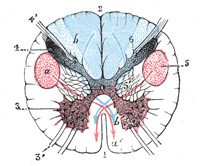 |
FIG. 687– Section of the medulla oblongata through the lower part of the decussation of the pyramids. (Testut.) 1. Anterior median fissure. 2. Posterior median sulcus. 3. Anterior column (in red), with 3’, anterior root. 4. Posterior column (in blue), with 4’, posterior roots. 5. Lateral cerebrospinal fasciculus. 6. Posterior funiculus. The red arrow, a, a’, indicates the course the lateral cerebrospinal fasciculus takes at the level of the decussation of the pyramids; the blue arrow, b, b’, indicates the course which the sensory fibers take. (See enlarged image) |
| |
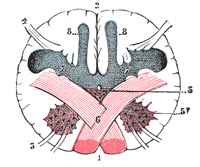 |
FIG. 688– Section of the medulla oblongata at the level of the decussation of the pyramids. (Testut.) 1. Anterior median fissure. 2. Posterior median sulcus. 3. Motor roots. 4. Sensory roots. 5. Base of the anterior column, from which the head (5’) has been detached by the lateral cerebrospinal fasciculus. 6. Decussation of the lateral cerebrospinal fasciculus. 7. Posterior columns (in blue). 8. Gracile nucleus. (See enlarged image) |
| |
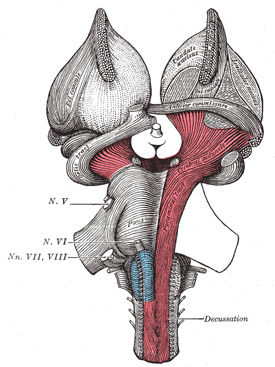 |
FIG. 689– Superficial dissection of brain-stem. Ventral view. (See enlarged image) |
| |
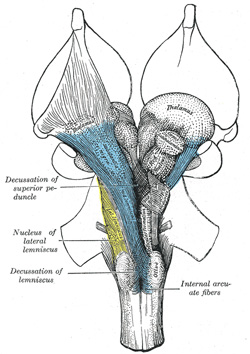 |
FIG. 690– Deep dissection of brain-stem. Ventral view. (See enlarged image) |
| |
| The fibers of the fasciculus gracilis and fasciculus cuneatus end by arborizing around the cells of these nuclei (Fig. 692). From the cells of the nuclei new fibers arise; some of these are continued as the posterior external arcuate fibers into the inferior peduncle, and through it to the cerebellum, but most of them pass forward through the neck of the posterior column, thus cutting off its head from its base (Fig. 693). Curving forward, they decussate in the middle line with the corresponding fibers of the opposite side, and run upward immediately behind the cerebrospinal fibers, as a flattened band, named the lemniscus or fillet. The decussation of these sensory fibers is situated above that of the motor fibers, and is named the decussation of the lemniscus or sensory decussation. The lemniscus is joined by the spinothalamic fasciculus (page 792), the fibers of which are derived from the cells of the gray substance of the opposite side of the medulla spinalis. | 19 |
| The base of the posterior column at first lies on the dorsal aspect of the central canal, but when the latter opens into the fourth ventricle, it appears in the lateral part of the rhomboid fossa. It forms the terminal nuclei of the sensory fibers of the vagus and glossopharyngeal nerves, and is associated with the vestibular part of the acoustic nerve and the sensory root of the facial nerve. Still higher, it forms a mass of pigmented cells—the locus cæruleus—in which some of the sensory fibers of the trigeminal nerve appear to end. The head of the posterior column forms a long nucleus, in which the fibers of the spinal tract of the trigeminal nerve largely end. | 20 |
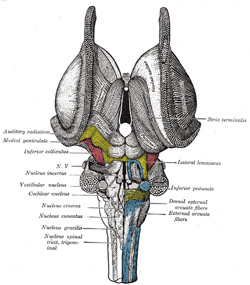 |
FIG. 691– Dissection of brain-stem. Dorsal view. The nuclear masses of the medulla are taken from model by Weed, Carnegie Publication, No. 19. (See enlarged image) |
| |
| The dorsal spinocerebellar fasciculus (fasciculus cerebellospinalis; direct cerebellar tract) leaves the lateral district of the medulla oblongata; most of its fibers are carried backward into the inferior peduncle of the same side, and through it are conveyed to the cerebellum; but some run upward with the fibers of the lemniscus, and, reaching the inferior colliculus, undergo decussation, and are carried to the cerebellum through the superior peduncle. | 21 |
| The proper fasciculi (basis bundles) of the anterior and lateral funiculi largely consist of intersegmental fibers, which link together the different segments of the medulla spinalis; they assist in the production of the formatio reticularis of the medulla oblongata, and many of them are accumulated into a fasciculus which runs up close to the median raphé between the lemniscus and the rhomboid fossa; this strand is named the medial longitudinal fasciculus, and will be again referred to. | 22 |
| |
| Gray Substance of the Medulla Oblongata (Figs. 694, 695).—In addition to the gracile and cuneate nuclei, there are several other nuclei to be considered. Some of these are traceable from the gray substance of the medulla spinalis, while others are unrepresented in it. | 23 |
| 1. The hypoglossal nucleus is derived from the base of the anterior column; in the lower closed part of the medulla oblongata it is situated on the ventrolateral aspect of the central canal; but in the upper part it approaches the rhomboid fossa, where it lies close to the middle line, under an eminence named the trigonum hypoglossi (Fig. 709). Numerous fibers connect the two nuclei, both nuclei send long dendrons across the midline to the opposite nucleus; commissure fibers also connect them. The nucleus measures about 2 cm. in length, and consists of large multipolar nerve cells, similar to those in the anterior column of the spinal cord, whose axons constitute the roots of the hypoglossal nerve. These nerve roots leave the ventral side of the nucleus, pass forward between the white reticular formation and the gray reticular formation, some between the inferior olivary nucleus and the medial accessory olivary nucleus, and emerge from the antero-lateral sulcus. | 24 |
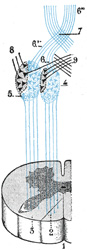 |
FIG. 692– Superior terminations of the posterior fasciculi of the medulla spinalis. 1. Posterior median sulcus. 2. Fasciculus gracilis. 3. Fasciculus cuneatus. 4. Gracile nucleus. 5. Cuneate nucleus. 6, 6’, 6’’. Sensory fibers forming the lemniscus. 7. Sensory decussation. 8. Cerebellar fibers uncrossed (in black). 9. Cerebellar fibers crossed (in black). (Testut.) (See enlarged image) |
| |
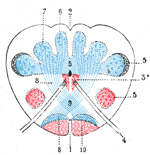 |
FIG. 693– Transverse section passing through the sensory decussation. (Schematic.) 1. Anterior median fissure. 2. Posterior median sulcus. 3, 3. Head and base of anterior column (in red). 4. Hypoglossal nerve. 5. Bases of posterior columns. 6. Gracile nucleus. 7. Cuneate nucleus. 8, 8. Lemniscus. 9. Sensory decussation. 10. Cerebrospinal fasciculus. (Testut.) (See enlarged image) |
| |
| 2. The nucleus ambiguus (Figs. 696, 697), the somatic motor nucleus of the glossopharyngeal, vagus and cranial portion of the accessory nerves, is the continuation into the medulla oblongata of the dorso-lateral cell group of the anterior column of the spinal cord. Its large multipolar cells are like those in the anterior column of the cord; they form a slender column in the deep part of the formatio reticularis grisea about midway between the dorsal accessory olive and the nucleus of the spinal tract of the trigeminal. It extends from the level of the decussation of the median fillet to the upper end of the medulla oblongata. Its fibers first pass backward toward the floor of the fourth ventricle and then curve rather abruptly lateralward and ventrally to join the fibers from the dorsal nucleus. | 25 |
| 3. The dorsal nucleus (Figs. 696, 698), nucleus ala cinerea, often called the sensory nucleus or the terminal nucleus of the sensory fibers of the glossopharyngeal and vagus nerves, is probably a mixed nucleus and contains not only the terminations of the sympathetic afferent or sensory fibers and the cells connected with them but contains also cells which give rise to sympathetic efferent or preganglionic fibers. These preganglionic fibers terminate in sympathetic ganglia from which the impulses are carried by other neurons. The cells of the dorsal nucleus are spindle-shaped, like those of the posterior column of the spinal cord, and the nucleus is usually considered as representing the base of the posterior column. It measures about 2 cm. in length, and in the lower, closed part of the medulla oblongata is situated behind the hypoglossal nucleus; whereas in the upper, open part it lies lateral to that nucleus, and corresponds to an eminence, named the ala cinerea (trigonum vagi), in the rhomboid fossa. | 26 |
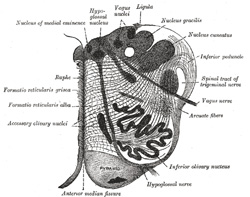 |
FIG. 694– Section of the medulla oblongata at about the middle of the olive. (Schwalbe.) (See enlarged image) |
| |
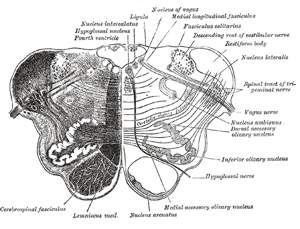 |
FIG. 695– Transverse section of medulla oblongata below the middle of the olive. (See enlarged image) |
| |
| 4. The nuclei of the cochlear and vestibular nerves are described on page 788. | 27 |
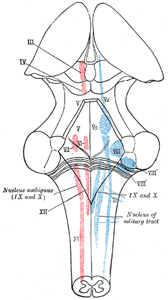 |
FIG. 696– The cranial nerve nuclei schematically represented; dorsal view. Motor nuclei in red; sensory in blue. (The olfactory and optic centers are not represented.) (See enlarged image) |
| |
| 5. The olivary nuclei (Fig. 694) are three in number on either side of the middle line, viz., the inferior olivary nucleus, and the medial and dorsal accessory olivary nuclei; they consist of small, round, yellowish cells and numerous fine nerve fibers. (a) The inferior olivary nucleus is the largest, and is situated within the olive. It consists of a gray folded lamina arranged in the form of an incomplete capsule, opening medially by an aperture called the hilum emerging from the hilum are numerous fibers which collectively constitute the peduncle of the olive. The axons, olivocerebellar fibers, which leave the olivary nucleus pass out through the hilum and decussate with those from the opposite olive in the raphé, then as internal arcuate fibers they pass partly through and partly around the opposite olive and enter the inferior peduncle to be distributed to the cerebellar hemisphere of the opposite side from which they arise. The fibers are smaller than the internal arcuate fibers connected with the median lemniscus. Fibers passing in the opposite direction from the cerebellum to the olivary nucleus are often described but their existence is doubtful. Much uncertainty also exists in regard to the connections of the olive and the spinal cord. Important connections between the cerebrum and the olive of the same side exist but the exact pathway is unknown. Many collaterals from the reticular formation and from the pyramids enter the inferior olivary nucleus. Removal of one cerebellar hemisphere is followed by atrophy of the opposite olivary nucleus. (b) The medial accessory olivary nucleus lies between the inferior olivary nucleus and the pyramid, and forms a curved lamina, the concavity of which is directed laterally. The fibers of the hypoglossal nerve, as they traverse the medulla, pass between the medial accessory and the inferior olivary nuclei. (c) The dorsal accessory olivary nucleus is the smallest, and appears on transverse section as a curved lamina behind the inferior olivary nucleus. | 28 |
| 6. The nucleus arcuatus is described below with the anterior external arcuate fibers. | 29 |
| |
| Inferior Peduncle (restiform body).—The position of the inferior peduncles has already been described (page 775). Each comprises: | 30 |
| (1) Fibers from the dorsal spinocerebellar fasciculus, which ascends from the lateral funiculus of the medulla spinalis. | 31 |
| (2) The olivocerebellar fibers from the opposite olivary nucleus. | 32 |
| (3) Internal arcuate fibers from the gracile and cuneate nuclei of the opposite side; these fibers form the deeper and larger part of the inferior peduncle. | 33 |
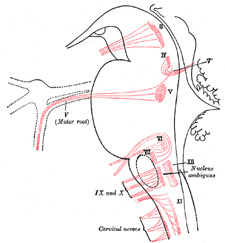 |
FIG. 697– Nuclei of origin of cranial motor nerves schematically represented; lateral view. (See enlarged image) |
| |
| (4) The anterior external arcuate fibers vary as to their prominence in different cases: in some they form an almost continuous layer covering the pyramid and olive, while in others they are barely visible on the surface. They arise from the cells of the gracile and cuneate nuclei, and passing forward through the formatio reticularis, decussate in the middle line. Most of them reach the surface by way of the anterior median fissure, and arch backward over the pyramid. Reinforced by others which emerge between the pyramid and olive, they pass backward over the olive and lateral district of the medulla oblongata, and enter the inferior peduncle. They thus connect the cerebellum with the gracile and cuneate nuclei of the opposite side. As the fibers arch across the pyramid, they enclose a small nucleus which lies in front of and medial to the pyramid. This is named the nucleus arcuatus, and is serially continuous above with the nuclei pontis in the pons; it contains small fusiform cells, around which some of the arcuate fibers end, and from which others arise. | 34 |
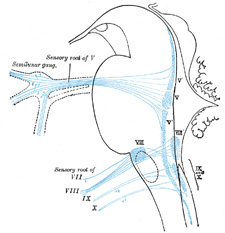 |
FIG. 698– Primary terminal nuclei of the afferent (sensory) cranial nerves schematically represented; lateral view. The olfactory and optic centers are not represented. (See enlarged image) |
| |
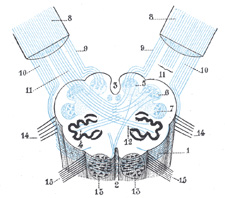 |
FIG. 699– Diagram showing the course of the arcuate fibers. (Testut.) 1. Medulla oblongata anterior surface. 2. Anterior median fissure. 3. Fourth ventricle. 4. Inferior olivary nucleus, with the accessory olivary nuclei. 5. Gracile nucleus. 6. Cuneate nucleus. 7. Trigeminal. 8. Inferior peduncles, seen from in front. 9. Posterior external arcuate fibers. 10. Anterior external arcuate fibers. 11. Internal arcuate fibers. 12. Peduncle of inferior olivary nucleus. 13. Nucleus arcuatus. 14. Vagus. 15. Hypoglossal. (See enlarged image) |
| |
| (5) The posterior external arcuate fibers also take origin in the gracile and cuneate nuclei; they pass to the inferior peduncle of the same side. It is uncertain whether fibers are continued directly from the gracile and cuneate fasciculi into the inferior peduncle. | 35 |
| (6) Fibers from the terminal sensory nuclei of the cranial nerves, especially the vestibular. Some of the fibers of the vestibular nerve are thought to continue directly into the cerebellum. | 36 |
| (7) Fibers from the ventral spinocerebellar fasciculus. | 37 |
| (8) The existence of fibers from the cerebellum (cerebellobulbar, cerebelloölivary, and cerebellospinal) to the medulla and spinal cord is very uncertain. | 38 |
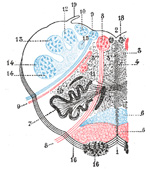 |
FIG. 700– The formatio reticularis of the medulla oblongata, shown by a transverse section passing through the middle of the olive. (Testut.) 1. Anterior median fissure. 2. Fourth ventricle. 3. Formatio reticularis, with 3’, its internal part (reticularis alba), and 3’’, its external part (reticularis grisea). 4. Raphé. 5. Pyramid. 6. Lemniscus. 7. Inferior olivary nucleus with the two accessory olivary nuclei. 8. Hypoglossal nerve, with 8’, its nucleus of origin. 9. Vagus nerve, with 9’, its nucleus of termination. 10. Lateral dorsal acoustic nucleus. 11. Nucleus ambiguus (nucleus of origin of motor fibers of glossopharyngeal, vagus, and cerebral portion of spinal accessory). 12. Gracile nucleus. 13. Cuneate nucleus. 14. Head of posterior column, with 14’, the lower sensory root of trigeminal nerve. 15. Fasciculus solitarius. 16. Anterior external arcuate fibers, with 16’, the nucleus arcuatus. 17. Nucleus lateralis 18. Nucleus of fasciculus teres. 19. Ligula. (See enlarged image) |
| |
| |
| Formatio Reticularis (Fig. 700).—This term is applied to the coarse reticulum which occupies the anterior and lateral districts of the medulla oblongata. It is situated behind the pyramid and olive, extending laterally as far as the inferior peduncles, and dorsally to within a short distance of the rhomboid fossa. The reticulum is caused by the intersection of bundles of fibers running at right angles to each other, some being longitudinal, others more or less transverse in direction. The formatio reticularis presents a different appearance in the anterior district from what it does in the lateral; in the former, there is an almost entire absence of nerve cells, and hence this part is known as the reticularis alba; whereas in the lateral district nerve cells are numerous, and as a consequence it presents a gray appearance, and is termed the reticularis grisea. | 39 |
| In the substance of the formatio reticularis are two small nuclei of gray matter: one, the inferior central nucleus (nucleus of Roller), near the dorsal aspect of the hilus of the inferior olivary nucleus; the other, the nucleus lateralis, between the olive and the spinal tract of the trigeminal nerve. | 40 |
| In the reticularis alba the longitudinal fibers form two well-defined fasciculi, viz.: (1) the lemniscus, which lies close to the raphé, immediately behind the fibers of the pyramid; and (2) the medial longitudinal fasciculus, which is continued upward from the anterior and lateral proper fasciculi of the medulla spinalis, and, in the upper part of the medulla oblongata, lies between the lemniscus and the gray substance of the rhomboid fossa. The longitudinal fibers in the reticularis grisea are derived from the lateral funiculus of the medulla spinalis after the lateral cerebrospinal fasciculus has passed over to the opposite side, and the dorsal spinocerebellar fasciculus has entered the inferior peduncle. They form indeterminate fibers, with the exception of a bundle named the fasciculus solitarius, which is made up of descending fibers of the vagus and glossopharyngeal nerves. The transverse fibers of the formatio reticularis are the arcuate fibers already described (page 782). | 41 |
| |
| The Pons (pons Varoli).—The pons or forepart of the hind-brain is situated in front of the cerebellum. From its superior surface the cerebral peduncles emerge, one on either side of the middle line. Curving around each peduncle, close to the upper surface of the pons, a thin white band, the tænia pontis, is frequently seen; it enters the cerebellum between the middle and superior peduncles. Behind and below, the pons is continuous with the medulla oblongata, but is separated from it in front by a furrow in which the abducent, facial, and acoustic nerves appear. | 42 |
| Its ventral or anterior surface (pars basilaris pontis) is very prominent, markedly convex from side to side, less so from above downward. It consists of transverse fibers arched like a bridge across the middle line, and gathered on either side into a compact mass which forms the middle peduncle. It rests upon the clivus of the sphenoidal bone, and is limited above and below by well-defined borders. In the middle line is the sulcus basilaris for the lodgement of the basilar artery; this sulcus is bounded on either side by an eminence caused by the descent of the cerebrospinal fibers through the substance of the pons. Outside these eminences, near the upper border of the pons, the trigeminal nerves make their exit, each consisting of a smaller, medial, motor root, and a larger, lateral, sensory root; vertical lines drawn immediately beyond the trigeminal nerves, may be taken as the boundaries between the ventral surface of the pons and the middle cerebellar peduncle. | 43 |
| Its dorsal or posterior surface (pars dorsalis pontis), triangular in shape, is hidden by the cerebellum, and is bounded laterally by the superior peduncle; it forms the upper part of the rhomboid fossa, with which it will be described. | 44 |
| |
| Structure (Fig. 701).—Transverse sections of the pons show it to be composed of two parts which differ in appearance and structure: thus, the basilar or ventral portion consists for the most part of fibers arranged in transverse and longitudinal bundles, together with a small amount of gray substance; while the dorsal tegmental portion is a continuation of the reticular formation of the medulla oblongata, and most of its constituents are continued into the tegmenta of the cerebral peduncles. | 45 |
| The basilar part of the pons consists of—(a) superficial and deep transverse fibers, (b) longitudinal fasciculi, and (c) some small nuclei of gray substance, termed the nuclei pontis which give rise to the transverse fibers. | 46 |
| The superficial transverse fibers (fibræ pontis superficiales) constitute a rather thick layer on the ventral surface of the pons, and are collected into a large rounded bundle on either side of the middle line. This bundle, with the addition of some transverse fibers from the deeper part of the pons, forms the greater part of the brachium pontis. | 47 |
| The deep transverse fibers (fibræ pontis profundæ) partly intersect and partly lie on the dorsal aspect of the cerebrospinal fibers. They course to the lateral border of the pons, and form part of the middle peduncle; the further connections of this brachium will be discussed with the anatomy of the cerebellum. | 48 |
| The longitudinal fasciculi (fasciculi longitudinales) are derived from the cerebral peduncles, and enter the upper surface of the pons. They stream downward on either side of the middle line in larger or smaller bundles, separated from each other by the deep transverse fibers; these longitudinal bundles cause a forward projection of the superficial transverse fibers, and thus give rise to the eminences on the anterior surface. Some of these fibers end in, or give off collateral to, the nuclei pontis. An important pathway is thus formed between the cerebral cortex and the cerebellum, the first neuron having its cell body in the cortex and sending its axon through the internal capsule and cerebral peduncle to form synapses either by terminals or collaterals with cell bodies situated in the nuclei pontis. Axons from these cells form the transverse fibers which pass through the middle peduncle into the cerebellum. Others after decussating, terminate either directly or indirectly in the motor nuclei of the trigeminal, abducent, facial, and hypoglossal nerves; but most of them are carried through the pons, and at its lower surface are collected into the pyramids of the medulla. The fibers which end in the motor nuclei of the cranial nerves are derived from the cells of the cerebral cortex, and bear the same relation to the motor cells of the cranial nerves that the cerebrospinal fibers bear to the motor cells in the anterior column of the medulla spinalis. Probably none of the collaterals or terminals of the cerebrospinal and cerebrobulbar fibers end directly in the motor nuclei of the spinal and cranial nerves, one or more association neurons are probably interpolated in the pathway. | 49 |
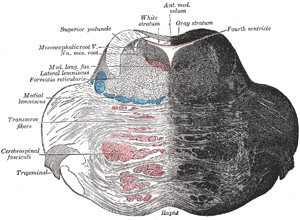 |
FIG. 701– Coronal section of the pons, at its upper part. (See enlarged image) |
| |
| The nuclei pontis are serially continuous with the arcuate nuclei in the medulla, and consist of small groups of multipolar nerve cells which are scattered between the bundles of transverse fibers. | 50 |
| The dorsal or tegmental part of the pons is chiefly composed of an upward continuation of the reticular formation and gray substance of the medulla oblongata. It consists of transverse and longitudinal fibers and also contains important gray nuclei, and is subdivided by a median raphé, which, however, does not extend into the basilar part, being obliterated by the transverse fibers. The transverse fibers in the lower part of the pons are collected into a distinct strand, named the trapezoid body. This consists of fibers which arise from the cells of the cochlear nucleus, and will be referred to in connection with the cochlear division of the acoustic nerve. In the substance of the trapezoid body is a collection of nerve cells, which constitutes the trapezoid nucleus. The longitudinal fibers, which are continuous with those of the medulla oblongata, are mostly collected into two fasciculi on either side. One of these lies between the trapezoid body and the reticular formation, and forms the upward prolongation of the lemniscus; the second is situated near the floor of the fourth ventricle, and is the medial longitudinal fasciculus. Other longitudinal fibers, more diffusely distributed, arise from the cells of the gray substance of the pons. | 51 |
| The rest of the dorsal part of the pons is a continuation upward of the formatio reticularis of the medulla oblongata, and, like it, presents the appearance of a network, in the meshes of which are numerous nerve cells. Besides these scattered nerve cells, there are some larger masses of gray substance, viz., the superior olivary nucleus and the nuclei of the trigeminal, abducent, facial, and acoustic nerves (Fig. 696). | 52 |
| 1. The superior olivary nucleus (nucleus olivaris superior) is a small mass of gray substance situated on the dorsal surface of the lateral part of the trapezoid body. Rudimentary in man, but well developed in certain animals, it exhibits the same structure as the inferior olivary nucleus, and is situated immediately above it. Some of the fibers of the trapezoid body end by arborizing around the cells of this nucleus, while others arise from these cells. | 53 |
| 2. The nuclei of the trigeminal nerve (nuclei n. trigemini) in the pons are two in number: a motor and a sensory. The motor nucleus is situated in the upper part of the pons, close to its posterior surface and along the line of the lateral margin of the fourth ventricle. It is serially homologous with the nucleus ambiguus and the dorso-lateral cell group of the anterior column of the spinal cord. The axis-cylinder processes of its cells form the motor root of the trigeminal nerve. The mesencephalic root arises from the gray substance of the floor of the cerebral aqueduct, joins the motor root and probably conveys fibers of muscle sense from the temporal, masseter and pterygoid muscles. It is not altogether clear whether the mesencephalic root is motor or sensory. The sensory nucleus is lateral to the motor one, and beneath the superior peduncle. Some of the sensory fibers of the trigeminal nerve end in this nucleus; but the greater number descend, under the name of the spinal tract of the trigeminal nerve, to end in the substantia gelatinosa of Rolando. The roots, motor and sensory, of the trigeminal nerve pass through the substance of the pons and emerge near the upper margin of its anterior surface. | 54 |
| 3. The nucleus of the abducent nerve (nucleus n. abducentis) is a circular mass of gray substance situated close to the floor of the fourth ventricle, above the striæ medullares and subjacent to the medial eminence: it lies a little lateral to the ascending part of the facial nerve. The fibers of the abducent nerve pass forward through the entire thickness of the pons on the medial side of the superior olivary nucleus, and between the lateral fasciculi of the cerebrospinal fibers, and emerge in the furrow between the lower border of the pons and the pyramid of the medulla oblongata. | 55 |
| 4. The nucleus of the facial nerve (nucleus n. fascialis) is situated deeply in the reticular formation of the pons, on the dorsal aspect of the superior olivary nucleus, and the roots of the nerve derived from it pursue a remarkably tortuous course in the substance of the pons. At first they pass backward and medialward until they reach the rhomboid fossa, close to the median sulcus, where they are collected into a round bundle; this passes upward and forward, producing an elevation, the colliculus facialis, in the rhomboid fossa, and then takes a sharp bend, and arches lateralward through the substance of the pons to emerge at its lower border in the interval between the olive and the inferior peduncle of the medulla oblongata. | 56 |
| 5. The nucleus of the cochlear nerve consists of: (a) the lateral cochlear nucleus, corresponding to the tuberculum acusticum on the dorso-lateral surface of the inferior peduncle; and (b) the ventral or accessory cochlear nucleus, placed between the two divisions of the nerve, on the ventral aspect of the inferior peduncle. | 57 |
| The nuclei of the vestibular nerve. (a) The medial (dorsal or chief vestibular nucleus), corresponding to the lower part of the area acustica in the rhomboid fossa; the caudal end of this nucleus is sometimes termed the descending or spinal vestibular nucleus. (b) The lateral or nucleus of Deiters, consisting of large cells and situated in the lateral angle of the rhomboid fossa; the dorso-lateral part of this nucleus is sometimes termed the nucleus of Bechterew. | 58 |
| The fibers of the vestibular nerve enter the medulla oblongata on the medial side of those of the cochlear, and pass between the inferior peduncle and the spinal tract of the trigeminal. They then divide into ascending and descending fibers. The latter end by arborizing around the cells of the medial nucleus, which is situated in the area acustica of the rhomboid fossa. The ascending fibers either end in the same manner or in the lateral nucleus, which is situated lateral to the area acustica and farther from the ventricular floor. Some of the axons of the cells of the lateral nucleus, and possibly also of the medial nucleus, are continued upward through the inferior peduncle to the roof nuclei of the opposite side of the cerebellum, to which also other fibers of the vestibular root are prolonged without interruption in the nuclei of the medulla oblongata. A second set of fibers from the medial and lateral nuclei end partly in the tegmentum, while the remainder ascend in the medial longitudinal fasciculus to arborize around the cells of the nuclei of the oculomotor nerve. | 59 |
| |
| The Cerebellum.—The cerebellum constitutes the largest part of the hindbrain. It lies behind the pons and medulla oblongata; between its central portion and these structures is the cavity of the fourth ventricle. It rests on the inferior occipital fossæ, while above it is the tentorium cerebelli, a fold of dura mater which separates it from the tentorial surface of the cerebrum. It is somewhat oval in form, but constricted medially and flattened from above downward, its greatest diameter being from side. Its surface is not convoluted like that of the cerebrum, but is traversed by numerous curved furrows or sulci, which vary in depth at different parts, and separate the laminæ of which it is composed. Its average weight in the male is about 150 gms. In the adult the proportion between the cerebellum and cerebrum is about 1 to 8, in the infant about 1 to 20. | 60 |
| |
| Lobes of the Cerebellum.—The cerebellum consists of three parts, a median and two lateral, which are continuous with each other, and are substantially the same in structure. The median portion is constricted, and is called the vermis, from its annulated appearance which it owes to the transverse ridges and furrows upon it; the lateral expanded portions are named the hemispheres. On the upper surface of the cerebellum the vermis is elevated above the level of the hemispheres, but on the under surface it is sunk almost out of sight in the bottom of a deep depression between them; this depression is called the vallecula cerebelli, and lodges the posterior part of the medulla oblongata. The part of the vermis on the upper surface of the cerebellum is named the superior vermis; that on the lower surface, the inferior vermis. The hemispheres are separated below and behind by a deep notch, the posterior cerebellar notch, and in front by a broader shallower notch, the anterior cerebellar notch. The anterior notch lies close to the pons and upper part of the medullas, and its superior edge encircles the inferior colliculi and the superior cerebellar peduncle. The posterior notch contains the upper part of the falx cerebelli, a fold of dura mater. | 61 |
| The cerebellum is characterized by a laminated or foliated appearance; it is marked by deep, somewhat curved fissures, which extend for a considerable distance into its substance, and divide it into a series of layers or leaves. The largest and deepest fissure is named the horizontal sulcus. It commences in front of the pons, and passes horizontally around the free margin of the hemisphere to the middle line behind, and divides the cerebellum into an upper and a lower portion. Several secondary but deep fissures separate the cerebellum into lobes, and these are further subdivided by shallower sulci, which separate the individual folia or laminæ from each other. Sections across the laminæ show that the folia, though differing in appearance from the convolutions of the cerebrum, are analogous to them, inasmuch as they consist of central white substance covered by gray substance. | 62 |
| The cerebellum is connected to the cerebrum, pons, and medulla oblongata; to the cerebrum by the superior peduncle, to the pons by the middle peduncle, and to the medulla oblongata by the inferior peduncles. | 63 |
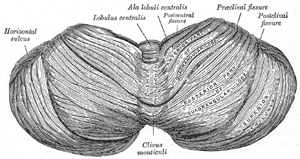 |
FIG. 702– Upper surface of the cerebellum. (Schäfer.) (See enlarged image) |
| |
| The upper surface of the cerebellum (Fig. 702) is elevated in the middle and sloped toward the circumference, the hemispheres being connected together by the superior vermis, which assumes the form of a raised median ridge, most prominent in front, but not sharply defined from the hemispheres. The superior vermis is subdivided from before backward into the lingula, the lobulus centralis, the monticulus and the folium vermis, and each of these, with the exception of the lingula, is continuous with the corresponding parts of the hemispheres—the lobulus centralis with the alæ, the monticulus with the quadrangular lobules, and the folium vermis with the superior semilunar lobules. | 64 |
| The lingula (lingula cerebelli) is a small tongue-shaped process, consisting of four or five folia; it lies in front of the lobulus centralis, and is concealed by it. Anteriorly, it rests on the dorsal surface of the anterior medullary velum, and its white substance is continuous with that of the velum. | 65 |
| The Lobulus Centralis and Alæ.—The lobulus centralis is a small square lobule, situated in the anterior cerebellar notch. It overlaps the lingula, from which it is separated by the precentral fissure; laterally, it extends along the upper and anterior part of each hemisphere, where it forms a wing-like prolongation, the ala lobuli centralis. | 66 |
| The Monticulus and Quadrangular Lobules.—The monticulus is the largest part of the superior vermis. Anteriorly, it overlaps the lobulus centralis, from which it is separated by the postcentral fissure; laterally, it is continuous with the quadrangular lobule in the hemispheres. It is divided by the preclival fissure into an anterior, raised part, the culmen or summit, and a posterior sloped part, the clivus; the quadrangular lobule is similarly divided. The culmen and the anterior parts of the quadrangular lobules form the lobus culminis; the clivus and the posterior parts, the lobus clivi. | 67 |
| |
| The Folium Vermis and Superior Semilunar Lobule.—The folium vermis (folium cacuminis; cacuminal lobe) is a short, narrow, concealed band at the posterior extremity of the vermis, consisting apparently of a single folium, but in reality marked on its upper and under surfaces by secondary fissures. Laterally, it expands in either hemisphere into a considerable lobule, the superior semilunar lobule (lobulus semilunaris superior; postero-superior lobules), which occupies the posterior third of the upper surface of the hemisphere, and is bounded below by the horizontal sulcus. The superior semilunar lobules and the folium vermis form the lobus semilunaris. | 68 |
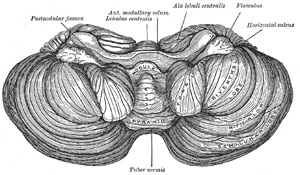 |
FIG. 703– Under surface of the cerebellum. (Schäfer.) (See enlarged image) |
| |
| The under surface of the cerebellum (Fig. 703) presents, in the middle line, the inferior vermis, buried in the vallecula, and separated from the hemisphere on either side by a deep groove, the sulcus valleculæ. Here, as on the upper surface, there are deep fissures, dividing it into separate segments or lobules; but the arrangement is more complicated, and the relation of the segments of the vermis to those of the hemispheres is less clearly marked. The inferior vermis is subdivided from before backward, into (1) the nodule, (2) the uvula, (3) the pyramid, and (4) the tuber vermis; the corresponding parts on the hemispheres are (1) the flocculus, (2) the tonsilla cerebelli, (3) the biventral lobule, and (4) the inferior semilunar lobule. The three main fissures are (1) the postnodular fissure, which runs transversely across the vermis, between the nodule and the uvula. In the hemispheres this fissure passes in front of the tonsil, crosses between the flocculus in front and the biventral lobule behind, and joins the anterior end of the horizontal sulcus. (2) The prepyramidal fissure crosses the vermis between the uvula in front and the pyramid behind, then curves forward between the tonsil and the biventral lobe, to join the postnodular fissure. (3) The postpyramidal fissure passes across the vermis between the pyramid and the tuber vermis, and, in the hemispheres, courses behind the tonsil and biventral lobules, and then along the lateral border of the biventral lobule to the postnodular sulcus; in the hemisphere it forms the anterior boundary of the inferior semilunar lobule. | 69 |
| The Nodule and Flocculus.—The nodule (nodulus vermis; nodular lobe), or anterior end of the inferior vermis, abuts against the roof of the fourth ventricle, and can only be distinctly seen after the cerebellum has been separated from the medulla oblongata and pons. On either side of the nodule is a thin layer of white substance, named the posterior medullary velum. It is semilunar in form, its convex border being continuous with the white substance of the cerebellum; it extends on either side as far as the flocculus. The flocculus is a prominent, irregular lobule, situated in front of the biventral lobule, between it and the middle cerebellar peduncle. It is subdivided into a few small laminæ, and is connected to the inferior medullary velum by its central white core. The flocculi, together with the posterior medullary velum and nodule, constitute the lobus noduli. | 70 |
| |
| The Uvula and Tonsilla.—The uvula (uvula vermis; uvular lobe) forms a considerable portion of the inferior vermis; it is separated on either side from the tonsil by the sulcus valleculæ, at the bottom of which it is connected to the tonsil by a ridge of gray matter, indented on its surface by shallow furrows, and hence called the furrowed band. The tonsilla (tonsilla cerebelli; amygdaline nucleus) is a rounded mass, situated in the hemispheres. Each lies in a deep fossa, termed the bird’s nest (nidus avis), between the uvula and the biventral lobule. The uvula and tonsillæ form the lobus uvulæ. | 71 |
| The Pyramid and Biventral lobules constitute the lobus pyramidis. The pyramid is a conical projection, forming the largest prominence of the inferior vermis. It is separated from the hemispheres by the sulcus valleculæ, across which it is connected to the biventral lobule by an indistinct gray band, analogous to the furrowed band already described. The biventral lobule is triangular in shape; its apex points backward, and is joined by the gray band to the pyramid. The lateral border is separated from the inferior semilunar lobule by the postpyramidal fissure. The base is directed forward, and is on a line with the anterior border of the tonsil, and is separated from the flocculus by the postnodular fissure. | 72 |
| The Tuber Vermis (tuber valvulæ) and the Inferior Semilunar Lobule (lobulus semilunaris inferior; postero-superior lobule) collectively form the lobus tuberus (tuberæ lobe). The tuber vermis, the most posterior division of the inferior vermis, is of small size, and laterally spreads out into the large inferior semilunar lobules, which comprise at least two-thirds of the inferior surface of the hemisphere. | 73 |
| |
| Internal Structure of the Cerebellum.—The cerebellum consists of white and gray substance. | 74 |
| |
| White Substance.—If a sagittal section (Fig. 704) be made through either hemisphere, the interior will be found to consist of a central stem of white substance, in the middle of which is a gray mass, the dentate nucleus. From the surface of this central white stem a series of plates is prolonged; these are covered with gray substance and form the laminæ. In consequence of the main branches from the central stem dividing and subdividing, a characteristic appearance, named the arbor vitæ, is presented. If the sagittal section be made through the middle of the vermis, it will be found that the central stem divides into a vertical and a horizontal branch. The vertical branch passes upward to the culmen monticuli, where it subdivides freely, one of its ramifications passing forward and upward to the central lobule. The horizontal branch passes backward to the folium vermis, greatly diminished in size in consequence of having given off large secondary branches; one, from its upper surface, ascends to the clivus monticuli; the others descend, and enter the lobes in the inferior vermis, viz., the tuber vermis, the pyramid, the uvula, and the nodule. | 75 |
| The white substance of the cerebellum includes two sets of nerve fibers: (1) projection fibers, (2) fibræ propriæ. | 76 |
| |
| Projection Fibers.—The cerebellum is connected to the other parts of the brain by three large bundles of projection fibers, viz., to the cerebrum by the superior peduncle, to the pons by the middle peduncle, and to the medulla oblongata by the inferior peduncles (Fig. 705). | 77 |
| The superior cerebellar peduncles (brachia conjunctiva), two in number, emerge from the upper and medial part of the white substance of the hemispheres and are placed under cover of the upper part of the cerebellum. They are joined to each other across the middle line by the anterior medullary velum, and can be followed upward as far as the inferior colliculi, under which they disappear. Below, they form the upper lateral boundaries of the fourth ventricle, but as they ascend they converge on the dorsal aspect of the ventricle and thus assist in roofing it in. | 78 |
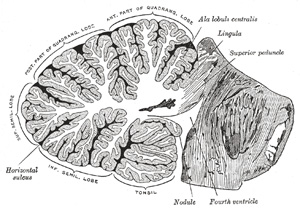 |
FIG. 704– Sagittal section of the cerebellum, near the junction of the vermis with the hemisphere. (Schäfer.) (See enlarged image) |
| |
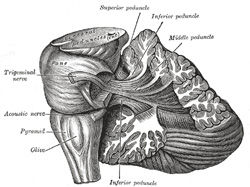 |
FIG. 705– Dissection showing the projection fibers of the cerebellum. (After E. B. Jamieson.) (See enlarged image) |
| |
| The fibers of the superior peduncle are mainly derived from the cells of the dentate nucleus of the cerebellum and emerge from the hilus of this nucleus; a few arise from the cells of the smaller gray nuclei in the cerebellar white substance, and others from the cells of the cerebellar cortex. They are continued upward beneath the corpora quadrigemina, and the fibers of the two peduncles undergo a complete decussation ventral to the Sylvian aqueduct. Having crossed the middle line they divide into ascending and descending groups of fibers, the former ending in the red nucleus, the thalamus, and the nucleus of the oculomotor nerve, while the descending fibers can be traced as far as the dorsal part of the pons; Cajal believes them to be continued into the anterior funiculus of the medulla spinalis. | 79 |
| As already stated (page 762), the majority of the fibers of the ventral spinocerebellar fasciculus of the medulla spinalis pass to the cerebellum, which they reach by way of the superior peduncle. | 80 |
| The middle cerebellar peduncles (brachia pontis) (Fig. 705) are composed entirely of centripetal fibers, which arise from the cells of the nuclei pontis of the opposite side and end in the cerebellar cortex; the fibers are arranged in three fasciculi, superior, inferior, and deep. The superior fasciculus, the most superficial, is derived from the upper transverse fibers of the pons; it is directed backward and lateralward superficial to the other two fasciculi, and is distributed mainly to the lobules on the inferior surface of the cerebellar hemisphere and to the parts of the superior surface adjoining the posterior and lateral margins. The inferior fasciculus is formed by the lowest transverse fibers of the pons; it passes under cover of the superior fasciculus and is continued downward and backward more or less parallel with it, to be distributed to the folia on the under surface close to the vermis. | 81 |
| The deep fasciculus comprises most of the deep transverse fibers of the pons. It is at first covered by the superior and inferior fasciculi, but crosses obliquely and appears on the medial side of the superior, from which it receives a bundle; its fibers spread out and pass to the upper anterior cerebellar folia. The fibers of this fasciculus cover those of the restiform body. 121 | 82 |
| The inferior cerebellar peduncles (restiform bodies) pass at first upward and lateralward, forming part of the lateral walls of the fourth ventricle, and then bend abruptly backward to enter the cerebellum between the superior and middle peduncles. Each contains the following fasciculi: (1) the dorsal spinocerebellar fasciculus of the medulla spinalis, which ends mainly in the superior vermis; (2) fibers from the gracile and cuneate nuclei of the same and of the opposite sides; (3) fibers from the opposite olivary nuclei; (4) crossed and uncrossed fibers from the reticular formation of the medulla oblongata; (5) vestibular fibers, derived partly from the vestibular division of the acoustic nerve and partly from the nuclei in which this division ends—these fibers occupy the medial segment of the inferior peduncle and divide into ascending and descending groups of fibers, the ascending fibers partly end in the roof nucleus of the opposite side of the cerebellum; (6) cerebellobulbar fibers which come from the opposite roof nucleus and probably from the dentate nucleus, and are said to end in the nucleus of Deiters and in the formatio reticularis of the medulla oblongata; (7) some fibers from the ventral spinocerebellar fasciculus are said to join the dorsal spinocerebellar fasciculus. | 83 |
| The anterior medullary velum (velum medullare anterius; valve of Vieussens; superior medullary velum) is a thin, transparent lamina of white substance, which stretches between the superior peduncle; on the dorsal surface of its lower half the folia and lingula are prolonged. It forms, together with the superior peduncle, the roof of the upper part of the fourth ventricle; it is narrow above, where it passes beneath the inferior colliculi, and broader below, where it is continuous with the white substance of the superior vermis. A slightly elevated ridge, the frænulum veli, descends upon its upper part from between the inferior colliculi, and on either side of this the trochlear nerve emerges. | 84 |
| The posterior medullary velum (velum medullare posterius; inferior medullary velum) is a thin layer of white substance, prolonged from the white center of the cerebellum, above and on either side of the nodule; it forms a part of the roof of the fourth ventricle. Somewhat semilunar in shape, its convex edge is continuous with the white substance of the cerebellum, while its thin concave margin is apparently free; in reality, however, it is continuous with the epithelium of the ventricle, which is prolonged downward from the posterior medullary velum to the ligulæ. | 85 |
| The two medullary vela are in contact with each other along their line of emergence from the white substance of the cerebellum; and this line of contact forms the summit of the roof of the fourth ventricle, which, in a vertical section through the cavity, appears as a pointed angle. | 86 |
| The Fibræ Propriæ of the cerebellum are of two kinds: (1) commissural fibers, which cross the middle line at the anterior and posterior parts of the vermis and connect the opposite halves of the cerebellum; (2) arcuate or association fibers, which connect adjacent laminæ with each other. | 87 |
| |
| Gray Substance.—The gray substance of the cerebellum is found in two situations: (1) on the surface, forming the cortex; (2) as independent masses in the anterior. | 88 |
| (1) The gray substance of the cortex presents a characteristic foliated appearance, due to the series of laminæ which are given off from the central white substance; these in their turn give off secondary laminæ, which are covered by gray substance. Externally, the cortex is covered by pia mater; internally is the medullary center, consisting mainly of nerve fibers. | 89 |
| |
| Microscopic Appearance of the Cortex (Fig. 706).—The cortex consists of two layers, viz., an external gray molecular layer, and an internal rust-colored nuclear layer; between these is an incomplete stratum of cells which are characteristic of the cerebellum, viz., the cells of Purkinje. | 90 |
| The external gray or molecular layer consists of fibers and cells. The nerve fibers are delicate fibrillæ, and are derived from the following sources: (a) the dendrites and axon collaterals of Purkinje’s cells; (b) fibers from cells in the nuclear layer; (c) fibers from the central white substance of the cerebellum; (d) fibers derived from cells in the molecular layer itself. In addition to these are other fibers, which have a vertical direction, and are the processes of large neuroglia cells, situated in the nuclear layer. They pass outward to the periphery of the gray matter, where they expand into little conical enlargements which form a sort of limiting membrane beneath the pia mater, analogous to the membrana limitans interna in the retina, formed by the sustentacular fibers of Müller. | 91 |
| The cells of the molecular layer are small, and are arranged in two strata, an outer and an inner. They all possess branched axons; those of the inner layer are termed basket cells; they run for some distance parallel with the surface of the folium—giving off collaterals which pass in a vertical direction toward the bodies of Purkinje’s cells, around which they become enlarged, and form basket-like net-works. | 92 |
| The cells of Purkinje form a single stratum of large, flask-shaped cells at the junction of the molecular and nuclear layers, their bases resting against the latter; in fishes and reptiles they are arranged in several layers. The cells are flattened in a direction transverse to the long axis of the folium, and thus appear broad in sections carried across the folium, and fusiform in sections parallel to the long axis of the folium. From the neck of the flask one or more dendrites arise and pass into the molecular layer, where they subdivide and form an extremely rich arborescence, the various subdivisions of the dendrites being covered by lateral spinelike processes. This arborescence is not circular, but, like the cell, is flattened at right angles to the long axis of the folium; in other words, it does not resemble a round bush, but has been aptly compared by Obersteiner to the branches of a fruit tree trained against a trellis or a wall. Hence, in sections carried across the folium the arborescence is broad and expanded; whereas in those which are parallel to the long axis of the folium, the arborescence, like the cell itself, is seen in profile, and is limited to a narrow area. | 93 |
| From the bottom of the flask-shaped cell the axon arises; this passes through the nuclear layer, and, becoming medullated, is continued as a nerve fiber in the subjacent white substance. As this axon traverses the granular layer it gives off fine collaterals, some of which run back into the molecular layer. | 94 |
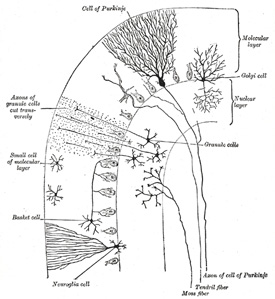 |
FIG. 706– Transverse section of a cerebellar folium. (Diagrammatic, after Cajal and Kölliker.) (See enlarged image) |
| |
| The internal rust-colored or nuclear layer (Fig. 706) is characterized by containing numerous small nerve cells of a reddish-brown color, together with many nerve fibrils. Most of the cells are nearly spherical and provided with short dendrites which spread out in a spider-like manner in the nuclear layer. Their axons pass outward into the molecular layer, and, bifurcating at right angles, run for some distance parallel with the surface. In the outer part of the nuclear layer are some larger cells, of the type II of Golgi. Their axons undergo frequent division as soon as they leave the nerve cells, and pass into the nuclear layer; while their dendrites ramify chiefly in the molecular layer. | 95 |
| Finally, in the gray substance of the cerebellar cortex there are fibers which come from the white center and penetrate the cortex. The cell-origin of these fibers is unknown, though it is believed that it is probably in the gray substance of the medulla spinalis. Some of these fibers end in the nuclear layer by dividing into numerous branches, on which are to be seen peculiar moss-like appendages; hence they have been termed by Ramón y Cajal the moss fibers; they form an arborescence around the cells of the nuclear layer and are said to come from fibers in the inferior peduncle. Other fibers, the clinging or tendril fibers, derived from the medullary center can be traced into the molecular layer, where their branches cling around the dendrites of Purkinje’s cells. They are said to come from fibers of the middle peduncle. | 96 |
| (2) The independent centers of gray substance in the cerebellum are four in number on either side: one is of large size, and is known as the nucleus dentatus; the other three, much smaller, are situated near the middle of the cerebellum, and are known as the nucleus emboliformis, nucleus globosus, and nucleus fastigii. | 97 |
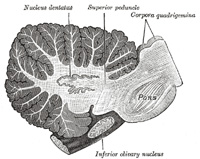 |
FIG. 707– Sagittal section through right cerebellar hemisphere. The right olive has also been cut sagitally. (See enlarged image) |
| |
| The nucleus dentatus (Fig. 707) is situated a little to the medial side of the center of the stem of the white substance of the hemisphere. It consists of an irregularly folded lamina, of a grayish-yellow color, containing white fibers, and presenting on its antero-medial aspect an opening, the hilus, from which most of the fibers of the superior peduncle emerge (page 792). | 98 |
| The nucleus emboliformis lies immediately to the medial side of the nucleus dentatus, and partly covering its hilus. The nucleus globosus is an elongated mass, directed antero-posteriorly, and placed medial to preceding. The nucleus fastigii is somewhat larger than the other two, and is situated close to the middle line at the anterior end of the superior vermis, and immediately over the roof of the fourth ventricle, from which it is separated by a thin layer of white substance. | 99 |
| The cerebellum is concerned with the coördination of movements necessary in equilibration, locomotion and prehension. In it terminate pathways conducting impulses of muscle sense, tendon sense, joint sense and equilibratory disturbances. With the exception of the ventral spinocerebellar fasciculus these impulses enter through the inferior peduncle. The reflex arc is completed by fibers in the superior peduncle which pass to the red nucleus and the thalamus and thence by additional neurons (rubrospinal tract) to the motor centers. The exact functions of its different parts are still quite uncertain, owing to the contradictory nature of the evidence furnished by (1) ablation experiments upon animals, and (2) clinical observations in man of the effects produced by abscesses or tumors affecting different portions of the organ. | 100 |
| |
| The Fourth Ventricle (ventriculus quartus).—The fourth ventricle, or cavity of the hind-brain, is situated in front of the cerebellum and behind the pons and upper half of the medulla oblongata. Developmentally considered, the fourth ventricle consists of three parts: a superior belonging to the isthmus rhombencephali, an intermediate, to the metencephalon, and an inferior, to the myelencephalon. It is lined by ciliated epithelium, and is continuous below with the central canal of the medulla oblongata; 122 above, it communicates, by means of a passage termed the cerebral aqueduct, with the cavity of the third ventricle. It presents four angles, and possesses a roof or dorsal wall, a floor or ventral wall, and lateral boundaries. | 101 |
| |
| Angles.—The superior angle is on a level with the upper border of the pons, and is continuous with the lower end of the cerebral aqueduct. The inferior angle is on a level with the lower end of the olive, and opens into the central canal of the medulla oblongata. Each lateral angle corresponds with the point of meeting of the brachia and inferior peduncle. A little below the lateral angles, on a level with the striæ medullares, the ventricular cavity is prolonged outward in the form of two narrow lateral recesses, one on either side; these are situated between the inferior peduncles and the flocculi, and reach as far as the attachments of the glossopharyngeal and vagus nerves. | 102 |
| |
| Lateral Boundaries.—The lower part of each lateral boundary is constituted by the clava, the fasciculus cuneatus, and the inferior peduncle; the upper part by the middle and the superior peduncle. | 103 |
| |
| Roof or Dorsal Wall (Fig. 708).—The upper portion of the roof is formed by the superior peduncle and the anterior medullary velum; the lower portion, by the posterior medullary velum, the epithelial lining of the ventricle covered by the tela chorioidea inferior, the tæniæ of the fourth ventricle, and the obex. | 104 |
| The superior peduncle (page 792), on emerging from the central white substance of the cerebellum, pass upward and forward, forming at first the lateral boundaries of the upper part of the cavity; on approaching the inferior colliculi, they converge, and their medial portions overlap the cavity and form part of its roof. | 105 |
| The anterior medullary velum (page 793) fills in the angular interval between the superior peduncle, and is continuous behind with the central white substance of the cerebellum; it is covered on its dorsal surface by the lingula of the superior vermis. | 106 |
| The posterior medullary velum (page 794) is continued downward and forward from the central white substance of the cerebellum in front of the nodule and tonsils, and ends inferiorly in a thin, concave, somewhat ragged margin. Below this margin the roof is devoid of nervous matter except in the immediate vicinity of the lower lateral boundaries of the ventricle, where two narrow white bands, the tæniæ of the fourth ventricle (ligulæ), appear; these bands meet over the inferior angle of the ventricle in a thin triangular lamina, the obex. The non-nervous part of the roof is formed by the epithelial lining of the ventricle, which is prolonged downward as a thin membrane, from the deep surface of the posterior medullary velum to the corresponding surface of the obex and tæniæ, and thence on to the floor of the ventricular cavity; it is covered and strengthened by a portion of the pia mater, which is named the tela chorioidea of the fourth ventricle. | 107 |
| The tæniæ of the fourth ventricle (tænia ventriculi quarti; ligula) are two narrow bands of white matter, one on either side, which complete the lower part of the roof of the cavity. Each consists of a vertical and a horizontal part. The vertical part is continuous below the obex with the clava, to which it is adherent by its lateral border. The horizontal portion extends transversely across the inferior peduncle, below the striæ medullares, and roofs in the lower and posterior part of the lateral recess; it is attached by its lower margin to the inferior peduncle, and partly encloses the choroid plexus, which, however, projects beyond it like a cluster of grapes; and hence this part of the tænia has been termed the cornucopia (Bochdalek). The obex is a thin, triangular, gray lamina, which roofs in the lower angle of the ventricle and is attached by its lateral margins to the clavæ. The tela chorioidea of the fourth ventricle is the name applied to the triangular fold of pia mater which is carried upward between the cerebellum and the medulla oblongata. It consists of two layers, which are continuous with each other in front, and are more or less adherent throughout. The posterior layer covers the antero-inferior surface of the cerebellum, while the anterior is applied to the structures which form the lower part of the roof of the ventricle, and is continuous inferiorly with the pia mater on the inferior peduncles and closed part of the medulla. | 108 |
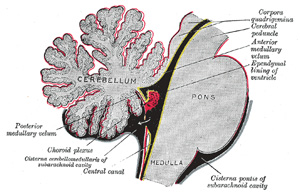 |
FIG. 708– Scheme of roof of fourth ventricle. The arrow is in the foramen of Majendie. (See enlarged image) |
| |
| |
| Choroid Plexuses.—These consist of two highly vascular inflexions of the tela chorioidea, which invaginate the lower part of the roof of the ventricle and are everywhere covered by the epithelial lining of the cavity. Each consists of a vertical and a horizontal portion: the former lies close to the middle line, and the latter passes into the lateral recess and projects beyond its apex. The vertical parts of the plexuses are distinct from each other, but the horizontal portions are joined in the middle line; and hence the entire structure presents the form of the letter T, the vertical limb of which, however, is double. | 109 |
| |
| Openings in the Roof.—In the roof of the fourth ventricle there are three openings, a medial and two lateral: the medial aperture (foramen Majendii), is situated immediately above the inferior angle of the ventricle; the lateral apertures, (foramina of Luschka are found at the extremities of the lateral recesses. By means of these three openings the ventricle communicates with the subarachnoid cavity, and the cerebrospinal fluid can circulate from the one to the other. | 110 |
| |
| Rhomboid Fossa (fossa rhomboidea; “floor” of the fourth ventricle) (Fig. 709).—The anterior part of the fourth ventricle is named, from its shape, the rhomboid fossa, and its anterior wall, formed by the back of the pons and medulla oblongata, constitutes the floor of the fourth ventricle. It is covered by a thin layer of gray substance continuous with that of the medulla spinalis; superficial to this is a thin lamina of neuroglia which constitutes the ependyma of the ventricle and supports a layer of ciliated epithelium. The fossa consists of three parts, superior, intermediate, and inferior. The superior part is triangular in shape and limited laterally by the superior cerebellar peduncle; its apex, directed upward, is continuous with the cerebral aqueduct; its base it represented by an imaginary line at the level of the upper ends of the superior foveæ. The intermediate part extends from this level to that of the horizontal portions of the tæniæ of the ventricle; it is narrow above where it is limited laterally by the middle peduncle, but widens below and is prolonged into the lateral recesses of the ventricle. The inferior part is triangular, and its downwardly directed apex, named the calamus scriptorius, is continuous with the central canal of the closed part of the medulla oblongata. | 111 |
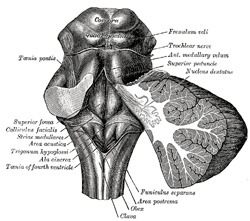 |
FIG. 709– Rhomboid fossa. (See enlarged image) |
| |
| The rhomboid fossa is divided into symmetrical halves by a median sulcus which reaches from the upper to the lower angles of the fossa and is deeper below than above. On either side of this sulcus is an elevation, the medial eminence, bounded laterally by a sulcus, the sulcus limitans. In the superior part of the fossa the medial eminence has a width equal to that of the corresponding half of the fossa, but opposite the superior fovea it forms an elongated swelling, the colliculus facialis, which overlies the nucleus of the abducent nerve, and is, in part at least, produced by the ascending portion of the root of the facial nerve. In the inferior part of the fossa the medial eminence assumes the form of a triangular area, the trigonum hypoglossi. When examined under water with a lens this trigone is seen to consist of a medial and a lateral area separated by a series of oblique furrows; the medial area corresponds with the upper part of the nucleus of the hypoglossal nerve, the lateral with a small nucleus, the nucleus intercalatus. | 112 |
| The sulcus limitans forms the lateral boundary of the medial eminence. In the superior part of the rhomboid fossa it corresponds with the lateral limit of the fossa and presents a bluish-gray area, the locus cæruleus, which owes its color to an underlying patch of deeply pigmented nerve cells, termed the substantia ferruginea. At the level of the colliculus facialis the sulcus limitans widens into a flattened depression, the superior fovea, and in the inferior part of the fossa appears as a distinct dimple, the inferior fovea. Lateral to the foveæ is a rounded elevation named the area acustica, which extends into the lateral recess and there forms a feebly marked swelling, the tuberculum acusticum. Winding around the inferior peduncle and crossing the area acustica and the medial eminence are a number of white strands, the striæ medullares, which form a portion of the cochlear division of the acoustic nerve and disappear into the median sulcus. Below the inferior fovea, and between the trigonum hypoglossi and the lower part of the area acustica is a triangular dark field, the ala cinerea, which corresponds to the sensory nucleus of the vagus and glossopharyngeal nerves. The lower end of the ala cinerea is crossed by a narrow translucent ridge, the funiculus separans, and between this funiculus and the clava, is a small tongue-shaped area, the area postrema. On section it is seen that the funiculus separans is formed by a strip of thickened ependyma, and the area postrema by loose, highly vascular, neuroglial tissue containing nerve cells of moderate size. | 113 |