| The early segmentation of the human ovum has not yet been observed, but judging from what is known to occur in other mammals it may be regarded as certain that the process starts immediately after the ovum has been fertilized, i. e., while the ovum is in the uterine tube. The segmentation nucleus exhibits the usual mitotic changes, and these are succeeded by a division of the ovum into two cells of nearly equal size. 5 The process is repeated again and again, so that the two cells are succeeded by four, eight, sixteen, thirty-two, and so on, with the result that a mass of cells is found within the zona striata, and to this mass the term morula is applied (Fig. 9). The segmentation of the mammalian ovum may not take place in the regular sequence of two, four, eight, etc., since one of the two first formed cells may subdivide more rapidly than the other, giving rise to a three-or a five-cell stage. The cells of the morula are at first closely aggregated, but soon they become arranged into an outer or peripheral layer, the trophoblast, which does not contribute to the formation of the embryo proper, and an inner cell-mass, from which the embryo is developed. Fluid collects between the trophoblast and the greater part of the inner cell-mass, and thus the morula is converted into a vesicle, the blastodermic vesicle (Fig. 10). The inner cell-mass remains in contact, however, with the trophoblast at one pole of the ovum; this is named the embryonic pole, since it indicates the situation where the future embryo will be developed. The cells of the trophoblast become differentiated into two strata: an outer, termed the syncytium or syncytiotrophoblast, so named because it consists of a layer of protoplasm studded with nuclei, but showing no evidence of subdivision into cells; and an inner layer, the cytotrophoblast or layer of Langhans, in which the cell outlines are defined. As already stated, the cells of the trophoblast do not contribute to the formation of the embryo proper; they form the ectoderm of the chorion and play an important part in the development of the placenta. On the deep surface of the inner cell-mass a layer of flattened cells, the entoderm, is differentiated and quickly assumes the form of a small sac, the yolk-sac. Spaces appear between the remaining cells of the mass (Fig. 11), and by the enlargement and coalescence of these spaces a cavity, termed the amniotic cavity (Fig. 12), is gradually developed. The floor of this cavity is formed by the embryonic disk composed of a layer of prismatic cells, the embryonic ectoderm, derived from the inner cell-mass and lying in apposition with the entoderm. | 1 |
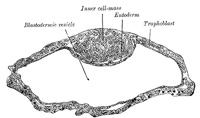 |
FIG. 10– Blastodermic vesicle of Vespertilio murinus. (After van Beneden.) (See enlarged image) |
| |
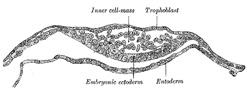 |
FIG. 11– Section through embryonic disk of Vespertilio murinus. (After van Beneden.) (See enlarged image) |
| |
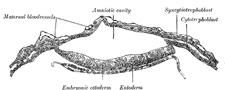 |
FIG. 12– Section through embryonic area of Vespertilio murinus to show the formation of the amniotic cavity. (After van Beneden.) (See enlarged image) |
| |
| |
| The Primitive Streak; Formation of the Mesoderm.—The embryonic disk becomes oval and then pear-shaped, the wider end being directed forward. Near the narrow, posterior end an opaque streak, the primitive streak (Figs. 13 and 14), makes its appearance and extends along the middle of the disk for about one-half of its length; at the anterior end of the streak there is a knob-like thickening termed Hensen’s knot. A shallow groove, the primitive groove, appears on the surface of the streak, and the anterior end of this groove communicates by means of an aperture, the blastophore, with the yolk-sac. The primitive streak is produced by a thickening of the axial part of the ectoderm, the cells of which multiply, grow downward, and blend with those of the subjacent entoderm (Fig. 15). From the sides of the primitive streak a third layer of cells, the mesoderm, extends lateralward between the ectoderm and entoderm; the caudal end of the primitive streak forms the cloacal membrane. | 2 |
 |
FIG. 13– Surface view of embryo of a rabbit. (After Kölliker.) arg. Embryonic disk. pr. Primitive streak. (See enlarged image) |
| |
| The extension of the mesoderm takes place throughout the whole of the embryonic and extra-embryonic areas of the ovum, except in certain regions. One of these is seen immediately in front of the neural tube. Here the mesoderm extends forward in the form of two crescentic masses, which meet in the middle line so as to enclose behind them an area which is devoid of mesoderm. Over this area the ectoderm and entoderm come into direct contact with each other and constitute a thin membrane, the buccopharyngeal membrane, which forms a septum between the primitive mouth and pharynx. In front of the buccopharyngeal area, where the lateral crescents of mesoderm fuse in the middle line, the pericardium is afterward developed, and this region is therefore designated the pericardial area. A second region where the mesoderm is absent, at least for a time, is that immediately in front of the pericardial area. This is termed the proamniotic area, and is the region where the proamnion is developed; in man, however, a proamnion is apparently never formed. A third region is at the hind end of the embryo where the ectoderm and entoderm come into apposition and form the cloacal membrane. | 3 |
| The blastoderm now consists of three layers, named from without inward: ectoderm, mesoderm, and entoderm; each has distinctive characteristics and gives rise to certain tissues of the body. 6 | 4 |
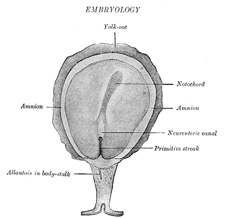 |
FIG. 14– Surface view of embryo of Hylobates concolor. (After Selenka.) The amnion has been opened to expose the embryonic disk. (See enlarged image) |
| |
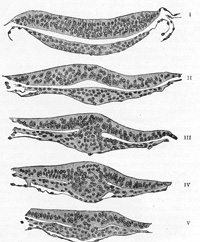 |
FIG. 15– Series of transverse sections through the embryonic disk of Tarsius. (After Hubrecht.) Section I passes through the disk, in front of Hensen’s knot and shows only the ectoderm and entoderm. Sections II, III, and IV pass through Hensen’s knot, which is seen in V tapering away into the primitive streak. In III, IV, and V the mesoderm is seen springing from the keel-like thickening of the ectoderm, which in III and IV is observed to be continuous into the entoderm. (See enlarged image) |
| |
| |
| Ectoderm.—The ectoderm consists of columnar cells, which are, however, somewhat flattened or cubical toward the margin of the embryonic disk. It forms the whole of the nervous system, the epidermis of the skin, the lining cells of the sebaceous, sudoriferous, and mammary glands, the hairs and nails, the epithelium of the nose and adjacent air sinuses, and that of the cheeks and roof of the mouth. From it also are derived the enamel of the teeth, and the anterior lobe of the hypophysis cerebri, the epithelium of the cornea, conjunctiva, and lacrimal glands, and the neuro-epithelium of the sense organs. | 5 |
| |
| Entoderm.—The entoderm consists at first of flattened cells, which subsequently become columnar. It forms the epithelial lining of the whole of the digestive tube excepting part of the mouth and pharynx and the terminal part of the rectum (which are lined by involutions of the ectoderm), the lining cells of all the glands which open into the digestive tube, including those of the liver and pancreas, the epithelium of the auditory tube and tympanic cavity, of the trachea, bronchi, and air cells of the lungs, of the urinary bladder and part of the urethra, and that which lines the follicles of the thyroid gland and thymus. | 6 |
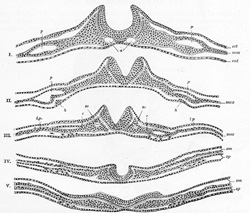 |
FIG. 16– A series of transverse sections through an embryo of the dog. (After Bonnet.) Section I is the most anterior. In V the neural plate is spread out nearly flat. The series shows the uprising of the neural folds to form the neural canal. a. Aortæ. c. Intermediate cell mass. ect. Ectoderm. ent. Entoderm. h, h. Rudiments of endothelial heart tubes. In III, IV, and V the scattered cells represented between the entoderm and splanchnic layer of mesoderm are the vasoformative cells which give origin in front, according to Bonnet, to the heart tubes, h; l.p. Lateral plate still undivided in I, II, and III; in IV and V split into somatic (sm) and splanchnic (sp) layers of mesoderm. mes. Mesoderm. p. Pericardium. so. Primitive segment. (See enlarged image) |
| |
| |
| Mesoderm.—The mesoderm consists of loosely arranged branched cells surrounded by a considerable amount of intercellular fluid. From it the remaining tissues of the body are developed. The endothelial lining of the heart and blood-vessels and the blood corpuscles are, however, regarded by some as being of entodermal origin. | 7 |
| As the mesoderm develops between the ectoderm and entoderm it is separated into lateral halves by the neural tube and notochord, presently to be described. A longitudinal groove appears on the dorsal surface of either half and divides it into a medial column, the paraxial mesoderm, lying on the side of the neural tube, and a lateral portion, the lateral mesoderm. The mesoderm in the floor of the groove connects the paraxial with the lateral mesoderm and is known as the intermediate cell-mass; in it the genito-urinary organs are developed. The lateral mesoderm splits into two layers, an outer or somatic, which becomes applied to the inner surface of the ectoderm, and with it forms the somatopleure; and an inner or splanchnic, which adheres to the entoderm, and with it forms the splanchnopleure (Fig. 16). The space between the two layers of the lateral mesoderm is termed the celom. | 8 |